Research 研究成果の紹介
花を咲かせる遺伝子の働きを切り替える仕組みを解明 促進する化学修飾を導入して活性化
~開花時期の操作や食料増産に期待~
花を咲かせる遺伝子の働きを切り替える仕組みを解明 促進する化学修飾を導入して活性化
~開花時期の操作や食料増産に期待~
【概要】
奈良先端科学技術大学院大学(学長:塩﨑一裕)先端科学技術研究科 バイオサイエンス領域の山口暢俊准教授・伊藤寿朗教授、東京工業大学の藤泰子准教授、中部大学の鈴木孝征教授、東京大学の稲垣宗一准教授・角谷徹仁教授らの共同研究グループは、植物の開花に関わる遺伝子の発現(注 1)を、DNA 分子が巻き付くヒストンというタンパク質の形の変化により、促進する仕組みを解明しました。この遺伝子の働きを抑制するポリコーム複合体(注 2)というタンパク質複合体を除去して、遺伝子発現の引き金になるヒストン修飾(注 3)を導入するものです。遺伝子は改変されないので、状況に応じて可逆的に変化させられます。ポリコーム複合体は動植物に保存されており、幅広い生物にこの仕組みが使われている可能性があります。この成果により、開花時期の調整など、園芸、農業に役立つことが期待されます。
本研究成果は、生命科学におけるオープンアクセス雑誌 e-Life 誌に 2024 年 9 月 10 日(火)午前11 時(英国夏時間)に公開されました(DOI:10.7554/eLife.100905.1)。
【背景と目的】
動物だけでなく、植物もその発生は受精卵という 1 つの細胞から始まります。その受精卵は細胞分裂と分化を繰り返して、様々な組織を構成する細胞種を作り出し、その体を作っていきます。同じゲノム DNA(注 4)を持つ細胞から異なる性質の細胞を生み出すうえで重要になるのは、ゲノム DNA の塩基配列は変化させずに遺伝子の発現を「オン」「オフ」どちらかにスイッチして制御するエピジェネティックな発現制御機構です。この遺伝子発現の制御機構を使えば、子孫に遺伝的な影響が出るゲノム DNA の塩基配列はそのまま保存し、必要に応じて可逆的に遺伝子発現の状況だけを変えて、望みの形質を作り出すことができることから、応用面での期待が高まっています。
ゲノム DNA の分子はヒストン(注 5)に巻きついたヌクレオソーム構造(注 6)をとっています。ヒストンが化学修飾を受けることで、遺伝子の発現が制御されるのです。例えばヒストンのメチル化は、ヒストンタンパク質の特定の残基にメチル基が付加される修飾で、遺伝子の発現を活性化または抑制する役割を持ちます。修飾が行われるアミノ酸残基やその状態によって、転写の抑制や促進が異なる影響を及ぼします。ヒストン修飾のうち、最もよく知られている例が、ヒストンを構成するタンパク質「H3」の 27 番目のリシン残基がトリメチル化される修飾(H3K27me3)です。この修飾は、ポリコーム複合体によって、発生や分化に重要な遺伝子に導入され、発現を抑制します。
モデル植物のシロイヌナズナ(注 7)では、発芽後しばらくは葉っぱだけを作り、それが光合成をして十分な栄養を蓄えます。その後、生長すると、花を作り、栄養を果実や種子に蓄えて、次の世代に子孫を残していきます。ポリコーム複合体は、花を作るのに必要な遺伝子に H3K27me3 を導入します。この修飾の導入により、遺伝子の発現を抑制して、花、果実、種子を早く作りすぎないようにしています。体に栄養があまりない状態で種子を作るより、栄養を十分蓄えて種子を作った方が、次の世代では生存に有利だと考えられるからです。実際に、ポリコーム複合体が働かない突然変異体では、体が十分に大きくならないまま、花が早く咲いてしまうことが知られています。これまでにポリコーム複合体が H3K27me3 を導入することは知られていましたが、どのようにしてその除去が行われるのかはわかっていませんでした。
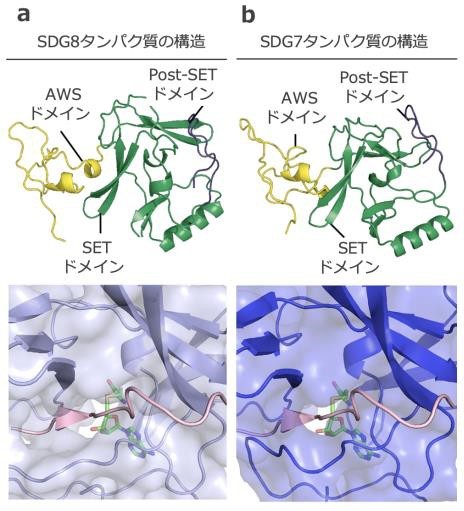
(a)SDG8 タンパク質にあるSET ドメインの構造.
(b)SDG7 タンパク質にあるSET ドメインの構造.
上部:AI を使った AWS ドメイン(黄色), SET ドメイン(緑色), Post-SET ドメイン(紫色)の構造予測. 下部:SDG とヒストン尾部(ピンク色)の相互作用. SDG タンパク質のポケット内にヒストン尾部が収まる様子がわかる.
【研究の内容】
ポリコーム複合体により抑制される遺伝子が、逆に促進されるようになる仕組みを知るために、本研究グループはヒストン H3 の 36 番目のリシン残基をトリメチル化される修飾(H3K36me3) が遺伝子の発現を促進することや、その修飾を導入する酵素に注目しました。この修飾は、SET DOMAIN GROUP(SDG)というヒストンメチル基転移酵素を含むタンパク質ファミリーが導入します。このファミリーのうち、SDG8 はH3K36me3 を導入することがすでに報告されていました。さらに SDG8 の主要な構造要素で触媒活性を担う部位の SET ドメインと非常によく似たドメインを SDG7 が持ち、ヒストン修飾が導入される部分とよく相互作用します(図 1)。 そこで、SDG7 と SDG8 という 2 つの酵素に注目して、ポリコーム複合体との関係を調べることにしました。
その結果、まずポリコーム複合体の構成因子(注 8)の 1 つである curly leaf(clf)変異体では、花を咲かせる遺伝子の発現を抑制することができません。そのため、この変異体では、花を咲かせる遺伝子など様々な遺伝子が発現する時期が早くなります。ところが、SDG7 と SDG8 遺伝子に変異を導入して、clf sdg7 sdg8 三重変異体を作ると、様々な遺伝子が発現する時期が一部回復して、すこし遅くなることがわかりました。このことは、ポリコーム複合体とSDG7やSDG8が逆の働きを持つことを意味します。
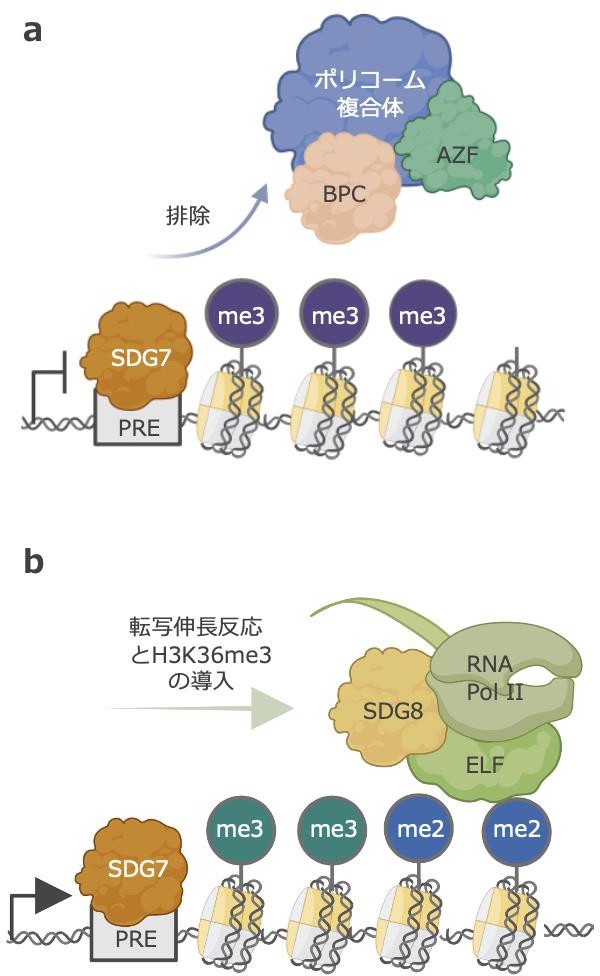
(a)SDG7 タンパク質によるポリコーム複合体の排除. 通常、H3K27me3 を導入するポリコーム複合体は, PRE に結合する. この配列にSDG7 が結合し, ポリコーム複合体を排除する.
(b)SDG8 タンパク質による転写伸長反応とH3K36me3 の導入. SDG7 がポリコーム複合体を排除した後, SDG8 が転写と H3K36me3 の導入をする.
PRE:ポリコーム結合配列. 紫:H3K27me3. 緑:H3K36me3
次に、ポリコーム複合体と SDG7 や SDG8 がどのようにして逆の働きをするのかを調べることにしました。ポリコーム複合体は、DNA 配列を認識して結合する AZFや BPC 転写因子(注 9)と相互作用して、主に遺伝子のプロモーター(注 10)に多く存在するポリコーム応答配列(注 11)を介して、ゲノム上に結合しています。そこで、SDG7 と SDG8 がゲノム上で結合している場所を調べ て、ポリコーム複合体がある位置と比べてみると、SDG7 は、ポリコーム複合体が位置するポリコーム応答配列付近に結合することがわかりました。さらに、SDG7 の機能を誘導する操作を行うと、花を咲かせる遺伝子などに結合しているポリコーム複合体と導入されていたH3K27me3 が排除されることがわかりました。
この実験結果から、SDG7 はポリコーム複合体とH3K27me3 を排除することで、花を咲かせる遺伝子の抑制を解除して、H3K36me3 を導入すると考えられます(図 2)。
一方で、SDG8 は遺伝子の内部に結合していることがわかりました。この SDG8 は真核生物の核内で mRNA の転写を担う酵素である RNA ポリメラーゼ II(注 12)や転写伸長因子(注 13) である ELF と相互作用することが報告されています。実際に、SDG8 とRNA ポリメラーゼII のゲノム上の結合部位はよく似ていることを明らかにしました。このことから、SDG8 は RNA ポリメラーゼと相互作用し、転写伸長反応と共役して、花を咲かせる遺伝子の内部に H3K36me3 を導入すると考えられます(図2)。
【今後の展開】
今回、花を咲かせないようにするヒストンの化学修飾が開花の方向に変わる仕組みの解明に成功しました。この仕組みをうまく使えば、ゲノム DNA を操作する方法では実現が困難とされた、開花の時期の可逆的なコントロールが可能になると考えられます。今後、応用研究を展開することで、農業や園芸の分野に大きな貢献が期待できます。
【用語解説】
注 1 遺伝子発現:DNA の情報が RNA に転写され、最終的にタンパク質として機能するプロセスを指す。
注2ポリコーム複合体:遺伝子の転写を抑制するタンパク質複合体で、特にヒストンのメチル化によりクロマチン構造を制御する。
注3ヒストン修飾:ヒストンタンパク質に対する化学的修飾で、遺伝子の発現を制御する重要なエピジェネティックなメカニズムである。
注 4 ゲノム DNA:生物の全遺伝情報を保持するDNA で、細胞の核内に存在する。
注 5 ヒストン:DNA が巻きついてクロマチンを形成するタンパク質で、遺伝子の発現調節に関わる。
注 6 ヌクレオソーム:DNA がヒストンに巻きついて形成される構造単位で、クロマチンの基本単位である。
注 7 シロイヌナズナ:植物研究のモデル生物で、遺伝学やエピジェネティクスの研究に広く用いられる。
注 8 ポリコーム複合体の構成因子:ポリコーム複合体を構成する因子。主に 4 つの因子が知られる。
注 9 転写因子:遺伝子のプロモーター領域に結合し、転写を促進または抑制するタンパク質。
注 10 プロモーター:遺伝子の転写開始点に位置する DNA 配列で、転写因子が結合し、遺伝子発現を制御する。
注 11 ポリコーム応答配列:ポリコーム複合体が結合して遺伝子の転写を抑制する特定のDNA 配列。
注 12 RNA ポリメラーゼ II:DNA から mRNA を合成する際に中心的な役割を果たす酵素で、特にタンパク質をコードする遺伝子の転写を行う。
注 13 転写伸長因子:RNA ポリメラーゼがDNA を読み進めてmRNA を合成する過程で、転写の効率と精度を向上させるために機能するタンパク質。
【掲載論文】
タイトル:Arabidopsis SDG proteins mediate Polycomb removal and transcription-coupled H3K36 methylation for gene activation
著者:Yicong Wang, Masato Abe, Yuka Kadoya, Takeru Saiki, Kanae Imai, Xuejing Wang, Taiko Kim To, Soichi Inagaki, Takamasa Suzuki, Tetsuji Kakutani, Toshiro Ito and Nobutoshi Yamaguchi
掲 載 誌 :e-Life
DOI:10.7554/eLife.100905.1
【花発生分子遺伝学研究室】
研究室紹介ページ:https://bsw3.naist.jp/courses/courses112.html
研究室ホームページ:https://bsw3.naist.jp/ito/
(2024年09月12日掲載)
この記事をシェア ![]()