Research 研究成果の紹介
遺伝子の優劣関係を決める新たな仕組みを解明、メンデルの優性の法則の謎を100年ぶりに ~有用な遺伝子を発現させる育種技術への応用に期待~
遺伝子の優劣関係を決める新たな仕組みを解明、メンデルの優性の法則の謎を100年ぶりに ~有用な遺伝子を発現させる育種技術への応用に期待~
奈良先端科学技術大学院大学(学長:小笠原直毅)バイオサイエンス研究科の和田七夕子(わだゆうこ)助教、高山誠司客員教授(現東京大学大学院農学生命科学研究科教授)らの研究グループは、農研機構、東北大学、大阪教育大学、神戸大学との共同研究により、どちらか片方の親の遺伝子の性質だけが子に現れるというメンデルの「優性の法則」として知られる現象について、複雑な優劣関係を決定する新たな仕組みを世界で初めて明らかにしました。
和田助教のコメント
 優劣性はメンデルの名前とともに広く知られていますが、その機構は不明なところが多いと言えます。エンドウマメの丸としわの例のように、劣性遺伝子が機能を失っている例が有名であり、対立遺伝子の機能欠損により優劣性が生まれると考えられていました。アブラナ(在来ナタネ)の花粉因子の優劣性について、過去に当研究室において、劣性遺伝子は機能的であり、低分子RNAを介したエピジェネティック制御により優劣性が制御されるという新奇な機構を明らかにし、注目を集めていました。アブラナにおける花粉因子の優劣性は、過去に仕組みが明らかにされた組み合わせ以外にも複雑な優劣関係が存在しますが、今回の報告はアブラナの花粉因子の複雑な優劣性は全て、エピジェネティック制御により説明されることを示したものです。
優劣性はメンデルの名前とともに広く知られていますが、その機構は不明なところが多いと言えます。エンドウマメの丸としわの例のように、劣性遺伝子が機能を失っている例が有名であり、対立遺伝子の機能欠損により優劣性が生まれると考えられていました。アブラナ(在来ナタネ)の花粉因子の優劣性について、過去に当研究室において、劣性遺伝子は機能的であり、低分子RNAを介したエピジェネティック制御により優劣性が制御されるという新奇な機構を明らかにし、注目を集めていました。アブラナにおける花粉因子の優劣性は、過去に仕組みが明らかにされた組み合わせ以外にも複雑な優劣関係が存在しますが、今回の報告はアブラナの花粉因子の複雑な優劣性は全て、エピジェネティック制御により説明されることを示したものです。
およそ100年前に遺伝学者の間で優劣性の起源が論争の的となり、Dominance modifierという因子が生まれることで優劣関係ができるという説が否定されました。しかし今回の結果から低分子RNAがDominance modifierそのものであり、その進化により複雑な優劣性が生まれたと考えられます。自家不和合性を制御する花粉因子以外にも、動植物において非常に複雑な優劣性が存在する例が知られます。そのような例においても、本論文で提唱したモデルが適用可能か、興味が持たれます。
本研究は本学客員教授である高山先生が中心となって、東北大をはじめとする多くの機関との長年にわたる共同研究により得られた成果であり、共同研究させて頂いた方にこの場を借りて感謝申し上げます。また、研究を中心となって進めてくれた安田晋輔君や、アブラナの形質転換と交配など長期間にわたる実験を支えてくれた、多くの研究室メンバーにも感謝します。
【概要】
奈良先端科学技術大学院大学(学長:小笠原直毅)バイオサイエンス研究科の和田七夕子(わだゆうこ)助教、高山誠司客員教授(現東京大学大学院農学生命科学研究科教授)らの研究グループは、農研機構、東北大学、大阪教育大学、神戸大学との共同研究により、どちらか片方の親の遺伝子の性質だけが子に現れるというメンデルの「優性の法則」として知られる現象について、複雑な優劣関係を決定する新たな仕組みを世界で初めて明らかにしました。
親から子へと遺伝子が受け継がれる遺伝現象において、片方の親の遺伝子の性質のみが子に現れる場合が多く見られます。これはメンデルの「優性の法則」として古くから知られており、性質として現れる遺伝子を優性遺伝子、発現しない方を劣性遺伝子と呼びます。これまで劣性遺伝子は一般に機能を失っているために性質が現れないと考えられてきましたが、同研究グループは、優性の遺伝子から作られる小さな分子(低分子RNA)が、劣性の遺伝子の働きを阻害するという全く異なる仕組みを発見しました。さらに今回新たに、この低分子RNAを構成する塩基(核酸塩基)の配列が変化することによって、特定の遺伝子同士で複雑な優劣関係が生み出されることを明らかにしました。約100年前、遺伝子間の優劣性を決定する因子が進化する可能性について遺伝学者間で激しい論争がなされましたが、今回、その時に想定された仮説の因子が低分子RNAであり、それが進化することを証明しました。
今回の研究は、遺伝子の優劣関係を制御する新たな仕組みを明らかにしただけでなく、有用な遺伝子を働かせたり、有害な遺伝子の働きを抑えたりする技術へと結びつく可能性があり、植物育種への応用が期待できます。
本成果は、英科学誌Natureの植物専門オンライン姉妹誌、Nature Plants(1月号:英国時間12月22日(木)午後4時)に掲載されます(プレス解禁日時:日本時間 平成28年12月23日(金)午前1時)。
【掲載論文】
論文タイトル:A complex dominance hierarchy is controlled by polymorphism of small RNAs and their targets
(和訳:低分子RNAとその標的の配列多型性によって複雑な階層的優劣性が制御される)
著者:Shinsuke Yasuda1†, Yuko Wada1†, Tomohiro Kakizaki2†, Yoshiaki Tarutani1, Eiko Miura-Uno1, Kohji Murase1,
Sota Fujii1, Tomoya Hioki1, Taiki Shimoda1, Yoshinobu Takada3, Hiroshi Shiba1, Takeshi Takasaki-Yasuda4, Go
Suzuki5, Masao Watanabe3, Seiji Takayama1,6 (†These authors contributed equally to this work.)
所属:1奈良先端科学技術大学院大学, 2農研機構, 3東北大学, 4神戸大学, 5大阪教育大学, 6東京大学
論文掲載誌:Nature Plants(1月号)
DOI:http://dx.doi.org/10.1038/nplants.2016.206
naistar:http://hdl.handle.net/10061/11328(NAIST Academic Repository:naistar)
本研究の研究材料であるアブラナの花が、掲載号の表紙を飾りました。
http://www.nature.com/nplants/volumes/3/issues/1
また、掲載号のNews & Viewsにも取り上げられました。
http://www.nature.com/articles/nplants2016210
【解説】
子供は両親から一組ずつの遺伝子を受け取るため、父親と母親の両方の性質を持ち合わせることになりますが、いずれか一方の性質のみを示す場合が多く知られています。これはメンデルの「優性の法則」として古くから知られる現象です。各遺伝子は父親・母親由来のもので一対になっており、それぞれを対立遺伝子と呼んでいます。その対立遺伝子間で複雑な優劣関係が生じる場合も知られており、例えば、4種の対立遺伝子(S1、S2、S3、S4 )の間でS1 > S2 > S3 > S4という階層的な遺伝子の優劣関係が見られる場合、両親からS1型とS2型の遺伝子を受け継ぐと子供にはS1型の遺伝子の性質のみが現れます。しかし、S2型とS3型の遺伝子を受け継ぐと、今度はS2型の遺伝子のみの性質が現れます。このような複雑な優劣関係がどのように決まるのかは未解明でありました。
【実験の手法・結果】
アブラナ科植物の自家不和合性*1を制御する働きがあるSP11*2遺伝子は複数の対立遺伝子の種類(S1, S2,... Sn)が存在し、一例としてS44 > S60 > S40 > S29という複雑な優劣関係の階層が示されています。
この仕組みを解明するため、研究グループは、在来ナタネBrassica rapaを用いて、個体それぞれのSP11遺伝子の近傍の領域を調査し、24個の塩基で構成される低分子RNA(Smi2)が作られることを見出しました。その性質を調べたところ、優性のSP11遺伝子近傍から作られるSmi2は、劣性側のSP11遺伝子の発現を調節する領域に対してのみ高い類似性を示していました。
そこで、階層的に優性であるS60型のSmi2を、比較的に劣性であるS40型やS29型のSP11遺伝子を持つ個体に人為的に導入したところ、S40型やS29型のSP11遺伝子の発現調節領域がメチル化という修飾を受け、遺伝子の働きがカギのかかったように抑制され、阻害されていることがわかりました。
その一方で、より優性であるS44型や、自身であるS60型のSP11遺伝子の働きを抑えることはありませんでした。このような結果から、遺伝子近傍領域から作られる低分子RNAが、他方の遺伝子配列と類似していることで攻撃目標とし、複雑な優劣関係を制御するという新たな仕組みが証明されました。
【本研究の意義】
遺伝子に優劣関係が生じる原因については、古くから激しい議論が繰り広げられてきました。約100年前にイギリスの遺伝学者によって優劣関係を制御する架空の因子が進化する可能性が提唱されました。しかし、その後、優性遺伝子は機能を持っているのに対し劣性遺伝子が機能を失っているという説が広く支持されるに至り、そのような因子の存在自体が疑問視されていました。
今回我々が見出した低分子RNAは、過去に提唱された仮説の因子そのものであり、100年を経て半ば忘れ去られていた説がようやく立証されたと言えます。さらに、遺伝子が低分子RNAを獲得し優性遺伝子となっていく道筋も明らかとなりました。アブラナ科植物において発見したこの仕組みは、動植物に広く存在する可能性もあり、今後の研究の進展が期待されます。
また、本知見を応用することで遺伝子の働きを人為的に調節できるようになるため、有用な遺伝子を働かせ、有害な遺伝子を働かせなくする等、新たな植物育種技術としての発展も期待できます。
【用語解説】
*1 自家不和合性: 自分の花粉が雌しべに付着しても受精せず種子を作らない性質のこと
*2 SP11:花粉の表面に付着した小型のタンパク質で、個体毎に構造が異なる多型性を示します。この多様性により、受粉の際に自己認識がなされます。
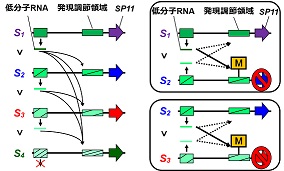
遺伝子間の優劣関係のモデル図(図左)。より優性のSP11遺伝子付近から作られる低分子RNAは自分よりも劣性側のSP11遺伝子の発現調節領域を標的とします。例えば両親からS1型とS2型のSP11遺伝子を受け継いだ場合、S1型(優性)の低分子RNAがS2型(劣性)のSP11遺伝子の発現調節領域をメチル化し(M)、遺伝子の働きを抑制します(図右上)。両親からS2型とS3型のSP11遺伝子を受け継いだ場合は、S2型の低分子RNAがS3型(さらに劣性)のSP11遺伝子発現調節領域をメチル化し、遺伝子の働きを抑制します(図右下)。
研究室紹介ページ:http://bsw3.naist.jp/courses/courses112.html
研究室ホームページ:http://bsw3.naist.jp/ito/
(2017年02月28日掲載)
この記事をシェア ![]()