Research 研究成果の紹介
植物の活発な炭酸ガス吸収、高生産能力を支える仕組みを解明- カギの遺伝子、NARA12を発見、食料増産に期待 -
分化・形態形成学講座(横田明穂教授)の博士課程学生の西村健司、博士研究員の小川太郎、助教の蘆田弘樹のグループは、モデル植物のシロイヌナズナを用いて植物葉緑体の高生産能力を支えるタンパク質遺伝子を見出してNARA12と命名。この遺伝子から作られるタンパク質(RH39)を中心にした高生産維持機能の仕組みを解明しました。この成果は、7月27日の奈良新聞と日刊工業新聞、7月28日の日経産業新聞に記事として取り上げられました。
プレスリリース詳細( 大学HP http://www.naist.jp/ 内コンテンツ )

【写真】記者会見後の写真
向かって右から横田明穂教授、博士研究員の小川太郎さん、博士課程学生の西村健司さん、蘆田弘樹助教
横田教授のコメント
この研究は、2000年頃から始めた葉緑体での光合成タンパク質の大量合成機構の解明を目指した研究の成果の一つです。西村君の先輩の小川太郎君(現在グローバルCOEの博士研究員)が大量合成できない変異株のスクリーニング法を構築し、これまでに数多くの変異株が分離されています。西村君はこの手法によって4種類の変異株を見つけ、nara12変異株はその内の一つです。
蘆田助教のコメント
医療用タンパク質などの有用タンパク質を植物の葉緑体で高生産させる研究が国際的に活発に行われています。私たちの研究室でも医療用タンパク質を植物葉緑体で合成させる研究を進めています。しかしながら、しばしば有用タンパク質の生産量が低い問題が生じます。NARA12研究によって明らかになった植物葉緑体の大量タンパク質合成機構は、有用タンパク質を大量に合成させるために応用できると期待されます。
掲載論文
Nishimura, K., Ashida, H., Ogawa, T. and Yokota, A. (2010) A DEAD box protein is required for formation of a hidden break in Arabidopsis chloroplast 23S rRNA. Plant J.
(リンクをクリックすると PubMed のページに移ります )
研究の概要
植物の葉は太陽エネルギーをCO2の炭素Cに蓄え、その一部はデンプンなどの栄養源として私たち動物に提供しています。この反応を支えているのが葉の細胞内小器官、葉緑体です。その規模は地球上で年間1000億トンにも達します。葉緑体は動物や細菌などの細胞のタンパク質の濃度に比べて2倍もの高濃度のタンパク質を合成してその目的を果たしています。しかしこれまで、葉緑体のタンパク質合成装置がなぜそれほどまでに高濃度のタンパク質を合成することができるのか、不明でした。今回英国誌プラントジャーナル電子版として発表された論文はこの高濃度タンパク質合成に関与する重要な遺伝子の発見を報じたものです。
大学院生の西村健司君(横田研D3)は、核染色体遺伝子に変異を持つシロイヌナズナ種子プールから葉緑体内でももっとも高濃度に合成される光合成CO2固定酵素ルビスコを合成できなくなった変異株をスクリーニングし、野生株の20%程度しかルビスコを合成できない変異株を見出しました。この変異株の詳細な解析の結果、この変異は葉緑体リボソーム23S RNA (23S rRNA)の巻き戻し酵素(ヘリカーゼ)RH39の遺伝子の変異が原因で、その遺伝子をNARA12と命名しました。葉緑体タンパク質合成の場であるリボソームを構成する23S rRNAの成熟化が不完全な場合、葉緑体は正常なタンパク質合成をおこなえず、ルビスコ以外にも葉緑体で合成される光合成タンパク質の量も激減していました。私たちは、RH39と未同定のRNaseのはたらき協働作業によって、タンパク質合成工場であるリボソームの機能が正常に維持されることが光合成にとって重要であると考えています(図1)。
私たちはすでに、ある種の原始紅藻がスーパールビスコを使って過酷環境下でも活発に光合成していることを見出しています。この遺伝子の発見後、世界中でこの遺伝子の植物葉緑体での発現に向け競争が起きましたが、現在まで成功していません。今回のNARA12遺伝子の発見はこの成功に重要なきっかけとなると期待しています。
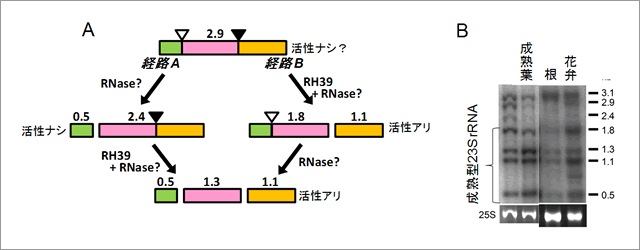
図1 葉緑体23SrRNAの成熟機構とNARA12(RH39)の働き
A : 本研究で明らかになった2.9kbの長さを持つ23SrRNA全長が成熟化を受けて活性型に変わる機構。全長23SrRNAは経路A及びBを経て切断を受け、機能型になる。経路Aではまず未知酵素で左末端が切断され、0.5kbと2.4kbが形成される。この2.4kbはまだ不活性型である。さらにNARA12から合成されるRH39が関わる切断によって2.4kbは1.3kbと1.1kbの活性型に変わる。経路Bでは、まずRH39が右側切断部位を切って1.8kbと1.1kbを形成する。この1.8kbも活性型であるが、その一部はさらに未知酵素で0.5kbと1.3kbに切断される。発見した変異株ではRH39が正常に機能できず、2.9kbと2.4kbが蓄積し、さらに大きな前駆体の蓄積も見られた。
B : シロイヌナズナ各組織の発達段階の異なる葉緑体(プラスチド)に含まれる23SrRNAの成熟度。根では大半が未成熟型(3.1kb)で成熟型はほとんど存在しない。花弁、さらには黄化子葉へとプラスチドの発達に応じて成熟化も進行していた。
(2010年08月05日掲載)
この記事をシェア ![]()