Research 研究成果の紹介
優性遺伝子が劣性遺伝子に勝つ新たな仕組みを解明
細胞間情報学講座の高山教授らのグループが、片方の親の形質のみが現れる「優劣性」の現象に、劣性側の対立遺伝子の発現を積極的に抑える新たな仕組みが存在することを明らかにしました。この研究成果は、英科学誌Nature(8月19日号)に掲載され、同号のNews & Viewsにも取り上げられました。また、読売新聞、日本経済新聞、産経新聞、日経産業新聞、日刊工業新聞、河北新報、朝日新聞に記事として掲載されました。
プレスリリース詳細( 大学HP http://www.naist.jp/ 内コンテンツ )
高山教授のコメント

高山教授の記者会見自家不和合性という多数の対立遺伝子を利用して自己・非自己を識別する仕組みを研究する過程で見出されてきた成果です。純系のモデル生物を扱うことが多い最近の研究では見落とされがちな「優劣性」現象ですが、多数の対立遺伝子を扱う自家不和合性研究では、対立遺伝子間に複雑な優劣性関係が存在することは古くから知られていました。また、従来知られている優劣性の仕組みが、劣性側対立遺伝子の機能欠損によるのに対し、自家不和合性の対立遺伝子は、すべて自己・非自己の識別という点で正常に機能することも分かっていましたので、従来とは異なる仕組みの関与が予測されました。そこで、実際に本現象を詳しく解析してみたところ、当初予想していた以上に意外な、古典的遺伝学とは無縁と考えていたエピジェネティックな遺伝子発現制御の関与が明らかにされてきたという次第です。最近、ヒトなどでも多くの遺伝子が片側対立遺伝子特異的な発現を示すことが明らかにされてきており、類似の機構が関与するのかどうか今後の研究の進展が期待されます。
掲載論文
研究の概要
子供は、両親から1組ずつ計2組の遺伝子セットを受け継ぐので、両親の性質を併せ持つことが予測される。しかし、メンデルが1世紀以上前に観察した様に、両親から受け継いだ性質の内、一方の性質のみが現れる場合も多く見られ、「優劣性」という言葉で説明されている。個々の対立遺伝子間の「優劣性」が、最終的な子孫の表現型を決定することになる。しかし、何故片方の親の性質のみが現れるのかについては未解明な部分が多く、これまでは、優性側の対立遺伝子が特定の機能を持つのに対し、劣性側の対立遺伝子が機能を欠失している例が報告されているに過ぎなかった。
今回我々は、アブラナ科植物の自家不和合性において認められる優劣性現象に着目して研究を行った。自家不和合性は、多くの植物が進化させてきた自殖を抑制するための仕組みであり、自己・非自己の識別は、多数の複対立遺伝子(S1, S2, ---, Sn)により制御されている。アブラナ科植物の場合、各S複対立遺伝子は、実際にはSP11およびSRKという密接に連鎖した2つの複対立遺伝子より構成されることが示されている。SP11は花粉表層のリガンド分子を、SRKは雌ずい側で機能するリセプター分子をコードしている。両分子ともに多型性に富むが、同一S遺伝子上のSP11とSRKが特異的に相互作用することで、自己の花粉の識別が行われ、不和合反応が起きることが明らかにされている(図1)。花粉表層のSP11リガンド分子は、葯(2n)で合成されるため、例えばS1S2というヘテロ体の葯から作られる花粉は、通常はS1とS2の両方の性質を併せ持ち(共優性)、S1またはS2を持つ雌ずいのいずれに対しても不和合性を示す。しかし、SP11複対立遺伝子の間には優劣性の関係が認めらる場合もあり、例えばS1がS2に対して優性の場合には、その花粉はS1のみの性質を示し、S2を持つ雌ずいに対して和合性となるなど、自他識別反応に変化が生じることが知られてきた。
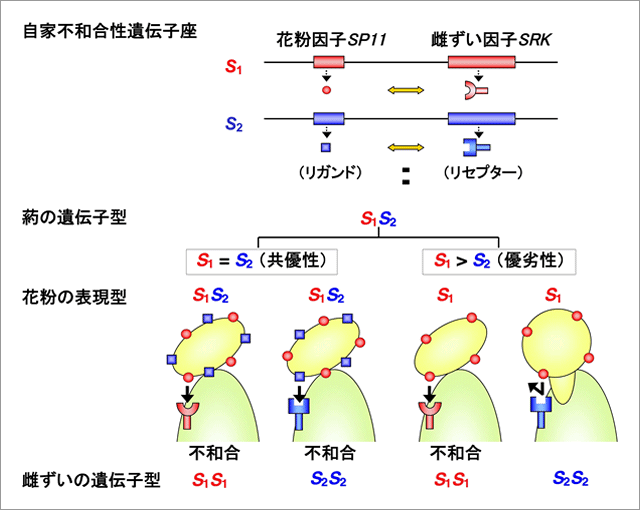
図1:アブラナ科植物の自家不和合性を支配するS複対立遺伝子と優劣性の関係
今回、我々はアブラナ科植物Brassica rapaを材料として、S8, S9, S12, S52などの複対立遺伝子が、S29, S40, S44, S60などの複対立遺伝子に対して優性となる仕組みについて解析した。その結果、SP11の発現部位である葯のタペート組織において、優性側のSP11遺伝子の隣接部位に存在する逆位反復配列から24ヌクレオチドからなる低分子RNAが作られること、それが引き金となって相同性を持つ劣性側のSP11遺伝子発現調節領域がメチル化を受け、発現が数万分の1にまで抑制されることが明らかとなった。優性側SP11遺伝子の発現調節領域には合成される低分子RNAと相同な配列がなく、優性側のSP11の発現が抑制されることはない。また、劣性側のSP11遺伝子の隣接部位(の逆位反復配列)からも相同な低分子RNAが合成されるが、それは1塩基置換により標的部位との相同性が低下しており、そのために自己のSP11に対してはメチル化を誘導しないことも明らかとなった(図2)。なお、今回劣性側対立遺伝子としてまとめて扱った対立遺伝子間にも、S44 > S60 > S40 > S29といったさらに複雑な優劣性関係が存在するが、その原因は未解明であり、研究を継続している。
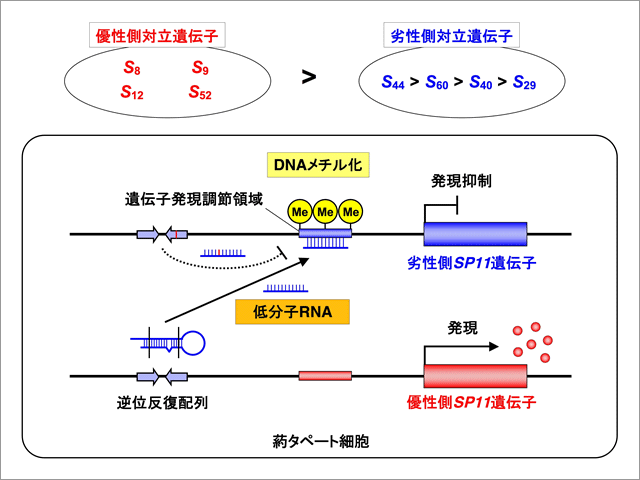
図2:自家不和合性の複対立遺伝子間における優劣性の分子機構モデル
本研究により、これまで優劣性という古典的な遺伝学の現象としてとらえられてきたものの中にも、エピジェネティックな遺伝子発現制御機構が関与する例が存在することが始めて明らかとなった。今回見出された機構が、他の優劣性現象や片側対立遺伝子特異的発現現象にどの程度関わっているか、今後さらに解析を進めていく必要がある。
なお、本研究は、生物系特定産業技術研究支援センターからのイノベーション創出基礎的研究推進事業、文部科学省および日本学術振興会からの科学研究費補助金の助成により成されたものである。
(2010年09月01日掲載)
この記事をシェア ![]()