Research 研究成果の紹介
植物は幹細胞の頑強な増殖を止めるために多段階スイッチを使っていた開花、種づくりに向けての巧妙な仕組み解明~ 園芸植物の花の改良や食糧増産に期待 ~
植物は幹細胞の頑強な増殖を止めるために多段階スイッチを使っていた
開花、種づくりに向けての巧妙な仕組み解明
~ 園芸植物の花の改良や食糧増産に期待 ~
【概要】
奈良先端科学技術大学院大学 (学長: 横矢直和) 先端科学技術研究科 バイオサイエンス領域の伊藤寿朗教授、中国南京大学のSun Bo(ソン・ボ)博士らと米国ペンシルバニア大の共同研究グループは、つぼみの中心部にある花の幹細胞※1が一定の数まで増殖したあと、使ってきた栄養を貯蔵するために増殖を止めるという重要な働きについて、その詳細な仕組みを世界に先駆けて明らかにしました。この成果により、花の大きさや数などが調節できるようになれば、園芸植物の花の改良や穀物類の増産などが期待できます。
花がおしべやめしべをつくり、次世代の種を残すためには、花の元となる幹細胞が十分に増殖した後に、それ以上の増殖を止めて、栄養を貯蔵する形に振り向ける必要があります。私たちはこれまでに幹細胞の増殖を助ける遺伝子の発現スイッチをオフにする複数の転写因子(タンパク質)※2の解析をおこなってきました。
遺伝子の本体である長いDNA※3はタンパク質であるヒストン※4に巻きつき、クロマチン※5と呼ばれるDNAが折りたたまれた構造を作ります。遺伝子発現がオンの時には、クロマチンは緩んだ構造をしており、逆に遺伝子発現のスイッチをオフにして、その状態を維持するためには、緩んでいるクロマチン構造をよりコンパクトに折りたたんだ状態に変えていかなければなりません。転写因子のひとつである「KNUCKLESタンパク質」は、幹細胞の増殖因子の発現をオフにすることはわかっていましたが、「どのような順序や方法でクロマチンに働きかけ、その構造と遺伝子の発現を変化させていくのか?」について詳しい仕組みは謎でした。
伊藤教授らの共同研究グループは、モデル植物のシロイヌナズナ※6を使って実験を重ねた結果、最初の段階でKNUCKLES転写因子が、クロマチンの構造を開いた状態に維持するのに必要な因子(クロマチンリモデリング因子)の結合を阻害し、幹細胞の増殖因子の発現抑制に働くことを見いだしました。
さらに、このKNUCKLES転写因子は、ポリコム因子複合体という遺伝子発現の抑制に関わる因子と直接結合して幹細胞増殖因子の遺伝子座に導入することで、クロマチン構造をコンパクトに閉じた状態にしていました。つまり、1つの転写因子が多段階スイッチとして働いていることがわかり、その順序や仕組みを突き止めることができました。
ポリコム因子※7がKNUCKLESと同じDNA結合領域を持つ転写因子により導入されるという報告はこれまでに動物でもなされています。しかし、幹細胞の増殖抑制を多段階に行う仕組みはこれまで動物において報告されておらず、植物の非常に強い幹細胞の活性を押さえ込むために必要であると考えられます。本研究は、植物の進化や生き残り戦略を知る上でも重要です。本研究の成果は2019年5月13日に米国の学会誌「The Plant Cell」に掲載されました。
伊藤寿朗教授のコメント

植物の幹細胞は動物と比べて、非常に強い増殖活性をもちます。私たちは植物において幹細胞の増殖抑制に働くスイッチの解析をしてきました。本論文は幹細胞の活性を直接抑制する重要因子が多段階のスイッチとして抑制の開始と抑制状態の維持を行っていることを世界に先駆けて発見しました。論文を書いてから発表されるまで時間がかかりましたが、その分とてもしっかりとした仕事になりました。 食糧増産のための基盤技術としてだけではなく、幹細胞の鍵因子の遺伝子発現抑制の詳細な仕組みを明らかに出来た意味合いは大きいと感じています。
【ご連絡事項】
(1)本件につきましては、奈良先端科学技術大学院大学から奈良県文化教育記者クラブをメインとし、学研都市記者クラブ、大阪科学・大学記者クラブに科学技術振興機構から文部科学記者会及び科学記者会に同時にご連絡しております。
(2)取材希望がございましたら、恐れ入りますが下記までご連絡願います。
(3)プレスリリースに関する問合せ先
奈良先端科学技術大学院大学 先端科学技術研究科 バイオサイエンス領域
花発生分子遺伝学研究室 伊藤 寿朗
TEL:0743-72-5500 FAX 0743-72-5502 E-mail: itot@bs.naist.jp
【背景】
植物は生殖器官であるおしべ、めしべをつくって、次の世代の子孫を残します。おしべ、めしべをつくった後に花の幹細胞は増殖を止めて、これまで自らが大きくなるために使っていた栄養を種に貯蔵するようになります。正しいタイミングで花幹細胞の増殖を止めることは、効率的に種子を形成するためにかかせないことです。これまでに幹細胞の増殖とその抑制に関わる転写因子については、複数あることはわかっていましたが、それらの転写因子がどのような順序や方法でクロマチンに働きかけて、その構造と遺伝子の発現を変化させていくのかについて、詳しい仕組みは未解明でした。
【研究手法と成果】
本研究グループは、まず、モデル植物であるシロイヌナズナを用い、幹細胞の増殖に必要な転写因子WUSCHEL(WUSタンパク質)とその上流の転写抑制因子KNUCKLES(KNUタンパク質)に注目しました。KNUタンパク質が作用しないknu突然変異体では、雌しべの中で種の代わりに過剰な数の花器官が出来て、幹細胞の増殖が止まらなくなることがわかっていたからです。また、KNUを過剰発現すると、逆に幹細胞の増殖が早くに止まってしまうため、花器官の数が減ります (図1)。そこで、knu突然変異体やKNU活性をコントロール出来る植物体を作成して、WUS遺伝子の発現とクロマチンに影響を及ぼすヒストン修飾 ※8という分子構造の変化を花発生の時間軸にそって解析しました。その結果、複数のヒストン修飾が時間軸に沿って変動し、時間の遅れが生じながらクロマチン構造が閉じていくことを発見しました。さらにKNUタンパク質に直接、相互作用するタンパク質として抑制的なヒストン修飾の導入に必要なポリコムタンパク質が含まれていました。
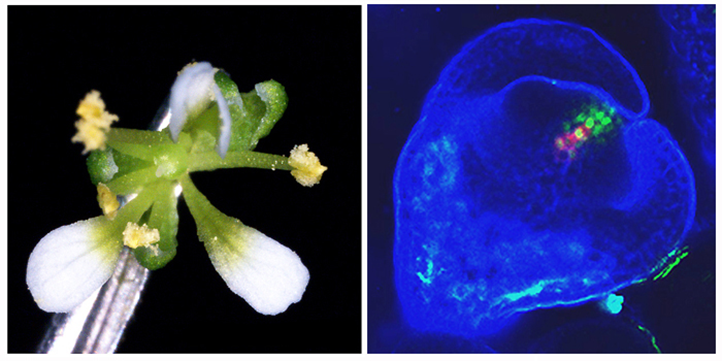
図1 図2
図1. 正しい花器官をつくるためには、花幹細胞が抑制される必要がある。KNUの過剰発現体では花器官の数が減っている。
図2. 若い花芽における幹細胞の増殖因子WUS(赤)と転写抑制因子KNU(緑)との発現パターン
ポリコムタンパク質によって導入される抑制的なヒストン修飾は遺伝子の発現をオフにする機能があるため、KNU誘導後のWUS遺伝子への発現の影響について調べたところ、抑制的ヒストン修飾が導入される前に、WUS遺伝子の発現量は減少することがわかりました。一方、ポリコム突然変異体の解析から、ポリコムは抑制し続けるために必要であることがわかりました。つまり、ポリコムは抑制の開始には関係しない一方、継続的な抑制状態の維持に必要であることがわかりました。それでは、どうやって、KNUはWUSの発現をオフにするのでしょうか?私たちは、KNUがWUS遺伝子座に直接結合して、クロマチンの構造を緩めることで遺伝子の活性化にかかわるSPLAYEDクロマチンリモデリングタンパク質の結合を阻害することを明らかにしました。この結果から、KNUタンパク質がWUS遺伝子に結合して多段階にはたらくスイッチとして、遺伝子の抑制に作用していることを明らかにしました。
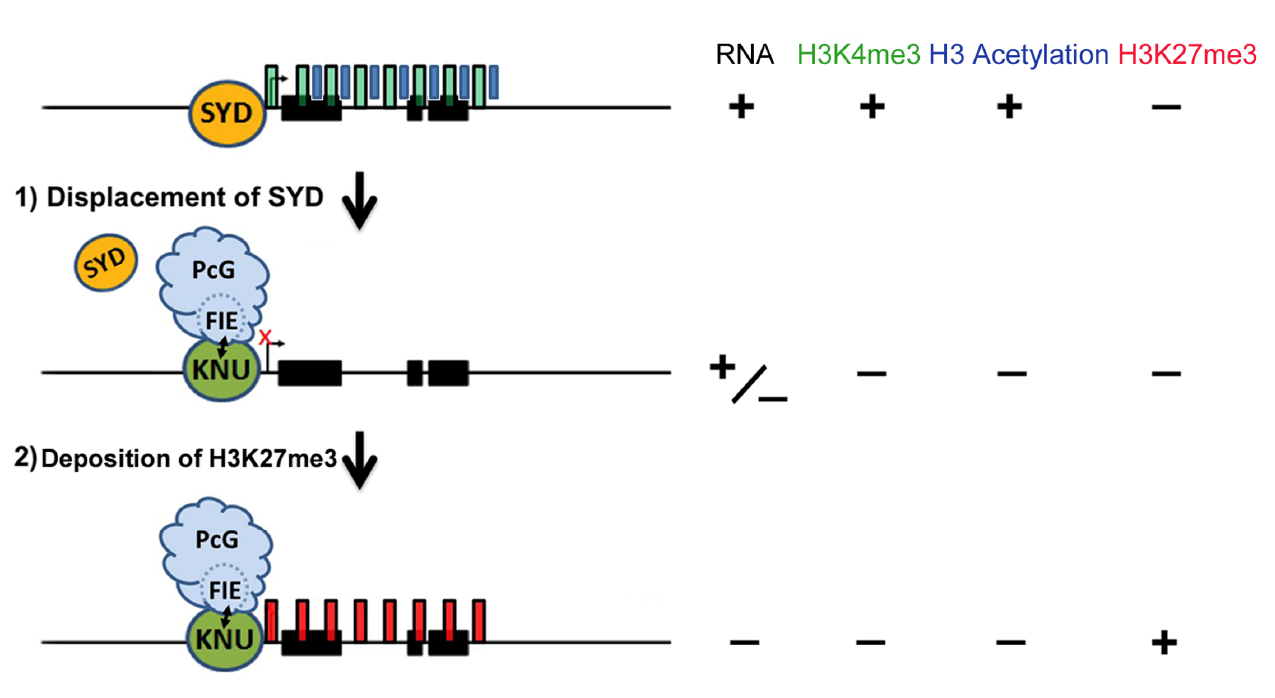
図3. WUS遺伝子の多段階的発現抑制機構
図3. WUS遺伝子のクロマチンはSYDクロマチンリモデリング因子により緩んだ構造が維持されている。一旦KNU転写因子が誘導されるとSYDを追い払い、WUS遺伝子の転写抑制が始まる。次にKNUタンパク質がポリコム因子(PcG)を導入することによって抑制的なヒストン修飾(H3K27me3)が付加されることにより、発現の抑制状態が維持される。
【波及効果】
植物の幹細胞は動物と比べて、非常に強い増殖活性をもちますが、その強い増殖能を花において抑制するくわしい仕組みはわかっていませんでした。本研究グループは植物において幹細胞の増殖抑制のために多段階のスイッチがあり抑制の開始と抑制状態の維持を行っていることを世界に先駆けて発見しました。また、転写因子の働きを変化させることで、めしべからつくられる果実や種子の大きさや数など様々な要因を人工的な調節で最適にできる可能性があり、食糧増産や安定供給が期待されます。
【掲載論文】
タイトル: Integration of transcriptional repression and Polycomb-mediated silencing of WUSCHEL in floral meristems
著者: Bo Suna,b,*, Yingying Zhoua, Jie Caia, Erlei Shanga, Nobutoshi Yamaguchic,d, Jun Xiaoe, Liang-Sheng Looib,c, Wan-Yi Weeb, Xiuying Gaoa, Doris Wagnere and Toshiro Itob,c,*
aState Key Laboratory of Pharmaceutical Biotechnology, School of Life Sciences, Nanjing University, Nanjing 210023, China; bTemasek Life Sciences Laboratory, 1 Research Link, National University of Singapore, 117604, Singapore; cBiological Sciences, Nara Institute of Science and Technology, 8916-5, Takayama, Ikoma, Nara, 630-0192, Japan; dPrecursory Research for Embryonic Science and Technology, Japan Science and Technology Agency, 4-1-8, Honcho, Kawaguchi-shi, Saitama, 332-0012, Japan; eDepartment of Biology, University of Pennsylvania, Philadelphia, PA 19104-6084, USA
掲載誌: The Plant Cell
【用語解説】
※1:幹細胞
多細胞体である動植物において、多種多様な細胞のもととなる細胞。分裂して自分と同じ細胞をつくる能力と別の種類の細胞に分化する能力をもっている。何千年も生き続ける縄文杉やジャイアントセコイヤに代表されるように、植物の幹細胞は動物と比べて非常に旺盛な増殖能力をもっている。
※2:転写因子
遺伝子の発現のオン、オフを切り替えるタンパク質。DNA上の特定の塩基配列に結合し、遺伝子の発現量を調節する。
※3:DNA
細胞の核内で長い鎖状の2重らせん構造を形成し、生物の遺伝情報を保持している物質。
※4:ヒストン
長いDNA を折りたたんで核内に収納するタンパク質。
※5:クロマチン
DNAをヒストンに巻きコンパクトに核内に収納する構造。遺伝子発現制御においても重要な役割を持っている。
※6:シロイヌナズナ
遺伝子の解析を行うのに適したアブラナ科の1年草。
※7:ポリコム因子
抑制的ヒストン修飾を付加する活性を持つタンパク質複合体
※8:ヒストン修飾
ヒストンタンパク質の特定のアミノ酸がメチル化修飾を受けることにより、遺伝子発現の活性化および、抑制が誘発される。ヒストンH3の27番目のリシン残基のメチル化(H3K27me3)は動物、植物でよく保存された抑制的なヒストン修飾マークである。
【原著論文のリンク】
http://www.plantcell.org/content/early/2019/05/08/tpc.18.00450
【花発生分子遺伝学研究室】
研究室紹介ページ:https://bsw3.naist.jp/courses/courses112.html
研究室ホームページ:https://bsw3.naist.jp/ito/
(2019年05月20日掲載)
この記事をシェア ![]()