Research 研究成果の紹介
持続可能資源である木質バイオマスの本体、 リグノセルロースの生合成モデルを25 年ぶりに刷新 ~ 産業利用の用途に即した質的改変技術の基盤開発へ ~
持続可能資源である木質バイオマスの本体、
リグノセルロースの生合成モデルを25 年ぶりに刷新
~ 産業利用の用途に即した質的改変技術の基盤開発へ ~
【概要】
奈良先端科学技術大学院大学(学長:横矢直和)先端科学技術研究科 バイオサイエンス領域 植物代謝制御研究室の出村拓教授、大谷美沙都助教らの研究グループはブリティッシュコロンビア大学(カナダ)との共同研究により、陸上で得られる最大の持続可能な次世代型資源として研究開発が進む木質バイオマス※1について、その本体である「リグノセルロース※2」という高分子混合物の作られる過程が従来の定説とは異なることをつきとめ、その生合成モデルを25年ぶりに刷新しました。リグノセルロースは、繊維やパルプなどに使われる「セルロース※3」、さまざまな糖原が得られる「ヘミセルロース※4」、香料や接着剤などの開発が進む「リグニン※5」の3成分で構成されます。これまで、最初に主成分のセルロースが蓄積され、それに依存して他の2成分が蓄積されるとなっていましたが、出村教授らは、セルロースが作られない変異体の解析から、この3成分は初期の段階でそれぞれ独立して蓄積されることを実証しました。産業利用の期待が高まる木質バイオマスは、成分の分離抽出が困難などの課題がありましたが、この成果により用途に応じた効果的な質的改変技術の基盤開発が期待されます。
リグノセルロースの生合成の詳細については、植物培養細胞から木質細胞※6への分化を誘導する実験系を使った研究や分子遺伝学的な研究によって大きく進展してきました。それらの成果から、これまでに、リグノセルロースの生合成の際には、最初にセルロースが木質細胞の細胞壁に堆積的に蓄積し、その後、ヘミセルロースおよびリグニンがセルロース依存的に細胞壁に蓄積する、というモデルが1990 年前後に提唱され、広く受け入れられてきました。
そこで、本研究では、培養細胞から木質細胞を分化させる実験により、セルロースを生合成するために必要な酵素の遺伝子が変化し、酵素の活性が失われた変異体「baculites※7(バキュリテス)」を新たに単離し、その詳細解析を行いました。その結果、木質細胞分化時にセルロース生合成が起こらないbaculites 変異体においても、ヘミセルロースやリグニンの細胞分化初期の蓄積が起こっていることを見出しました。さらに、ヘミセルロースとリグニンの蓄積パターンは細胞の骨格をなす繊維状の微小管※8のパターンに直接的に依存していること、細胞分化中期以降のヘミセルロースおよびリグニンの蓄積は、従来言われていたとおりセルロースの存在に大きく依存すること、も明らかにしました。これらの結果は、リグノセルロースの生合成初期では、セルロースとヘミセルロース、リグニンの蓄積は独立して制御されていることを示しており、従来のリグノセルロースの生合成モデルを大きく修正する成果です。
この成果は米国の植物生理学会の学会誌であるPlant Cell 誌オンラインサイトに日本時間平成30年10 月19 日(金)午前6 時付で掲載されました。
【解説】
陸上最大のバイオマス源は、陸上植物が蓄えているリグノセルロースを本体とする木質バイオマスです。リグノセルロースは、その大部分(約50%)を占めるセルロース、ヘミセルロース(20~30%)およびフェノール性ポリマー(高分子)であるリグニン(30~20%)からなっており、持続可能な次世代型資源として産業利用に向けた研究開発が進んでいます。その一つとして、目的に合わせた性質をもったリグノセルロースを生合成する試みがありますが、このためには、植物がどのようにリグノセルロースを作り出しているのかを理解することが重要です。リグノセルロース生合成過程の分子メカニズムについては、1980 年頃から急速に解明が進み、その成果として、「リグノセルロース生産細胞である木質細胞への分化が始まると、最初にセルロース蓄積が始まる」。次いで「セルロース蓄積とほぼ同時、あるいは少し遅れて、ヘミセルロースが蓄積し始める」。そして「最終的に時間をかけてリグニンが蓄積し、強固なリグノセルロースが完成する」というモデル(図1A)が提唱され、広く受け入れられてきました。今回、研究グループは、セルロース生合成が起こらなくてもヘミセルロースとリグニンは木質細胞の分化初期から蓄積すること、こうしたヘミセルロースとリグニンの蓄積は細胞骨格の制御を直接的に受けることを示しました。これによって、1)細胞分化の初期にはセルロースとヘミセルロースおよびリグニンの蓄積は独立して制御される。2)細胞分化の中期以降のヘミセルロースおよびリグニンの蓄積は、従来言われていたとおりセルロースの存在に大きく依存する、という新しいリグノセルロース生合成モデル(図1B)を提出することに成功しました。これは、従来のモデルを四半世紀ぶりに大きく修正する成果です。
【実験の手法・結果】
今回の研究では、木質細胞の一つで通水機能をもつ道管細胞を人為的に分化誘導できるシステムを導入したモデル植物・シロイヌナズナを材料にして、道管形成の過程を観察して新規変異体の単離と解析を行いました。新たに単離された変異体baculites は、通常、道管細胞分化誘導後に観察されるらせん状のリグノセルロース沈着が起こりません(図2)。解析の結果、baculites はセルロース生合成酵素遺伝子CESA7 の機能欠損型変異体であり、道管細胞の分化誘導後にセルロース生合成が全く起こらない一方で、ヘミセルロースの一種であるキシランや、リグニンといった非セルロース成分についてはらせん状に蓄積していることが示されました(図2)。さらに、細胞骨格である微小管の重合阻害剤で処理し、微小管がらせん構造を取れなくすると、キシランやリグニンのらせん状の蓄積は見られなくなり、乱れた微小管と一致する形で蓄積することも分かりました。
また、野生型シロイヌナズナでは、セルロースが蓄積するにしたがって、キシラン蓄積を示すシグナルが強く明瞭に線状化していくのに対して、baculites 変異体では、分化初期に見られる乱れたパッチ状のシグナルのまま、細胞の分化が完了していました(図2)。こうした結果から、リグノセルロース生合成におけるキシランとリグニンの蓄積過程はセルロース蓄積に非依存的な初期の段階と、蓄積したセルロースに依存する中期以降に大きく分けられること、さらに、キシランとリグニンの蓄積は、セルロースとは一部独立する形で微小管のパターンに依存していることを示唆しています。
【本研究の意義】
今回の研究により、従来のリグノセルロース生合成モデルが四半世紀ぶりに修正され、リグノセルロースの生合成メカニズムの分子的理解がさらに深まったといえます。これまでは従来のモデルにしたがって、リグノセルロース生合成の鍵はセルロースの生合成であり、その制御がヘミセルロースやリグニンの蓄積を支配すると考えられてきましたが、今回の成果によってこうした見方が大きく揺らぎ、セルロースと独立してヘミセルロースやリグニンを人為操作できる可能性が見出されました。これは、目的に合わせた性質をもったリグノセルロースの生合成を目指す上で、重要な発見です。本研究をさらに発展させることで、木質バイオマス中のセルロース、ヘミセルロース、リグニンの蓄積量比や蓄積箇所を自在にコントロールが可能になり、将来的な木質バイオマスの質的改変技術へつながることが期待されます。
【掲載論文】
タイトル:Patterned deposition of xylan and lignin is independent from the secondary wall cellulose of Arabidopsis xylem vessels
(和訳:シロイヌナズナ木部道管では、パターンをもったキシランとリグニンの沈着は二次壁セルロースとは独立である)
著者:Yuto Takenakaa, Yoichiro Watanabea,b, Mathias Schuetzb, Faride Undab, Joseph L Hill, Jrc, Pawittra Phookaewa, Arata Yonedaa, Shawn D Mansfieldb, Lacey Samuelsb, Misato Ohtania,* , and Taku Demura a,*
所属:a奈良先端科学技術大学院大学,bブリティッシュコロンビア大学,cペンシルベニア州立大学*:責任著者
掲載誌:Plant Cell
【用語解説】
※1 木質バイオマス:
バイオマスとは生物資源量のことであり、木材からなるバイオマスのことをとくに木質バイオマスとよぶ。木質バイオマスは樹木が光合成によって大気中の二酸化炭素を吸収・固定して作り出した炭素化合物であるため、持続可能かつ再生可能な資源であることから、化石燃料に変わる新しいエネルギー・材料資源として、利用技術の開発が盛んに行われている。
※2 リグノセルロース:
木質バイオマスの主体であり、セルロース、ヘミセルロースといった多糖性ポリマーとフェノール性ポリマーであるリグニンから構成される。リグノセルロースはこれら複数種の高分子ポリマーが複雑に絡み合った高次構造を形成しているため、難溶性の高分子混合物となっており、有用物質生産のために必要とされる各成分の抽出精製の難しさが、リグノセルロースの高度利用の妨げとなっている。
※3 セルロース:
多数のβ- D-グルコース分子が直鎖状に重合した天然高分子であり、植物細胞が有する細胞壁の主成分である。地球上で最も多く存在する炭水化物であり、古くから繊維として衣類や紙製品といった幅広い用途に利用されてきた。近年では、セルロースをナノレベルまで解きほぐしたナノセルロースを用いたナノセルロース化学が発展し、軽い・強い・固いといった特性を活かした新規素材の開発が加速している。多数のβ- D-グルコース分子が直鎖状に重合した天然高分子であり、植物細胞が有する細胞壁の主成分である。地球上で最も多く存在する炭水化物であり、古くから繊維として衣類や紙製品といった幅広い用途に利用されてきた。近年では、セルロースをナノレベルまで解きほぐしたナノセルロースを用いたナノセルロース化学が発展し、軽い・強い・固いといった特性を活かした新規素材の開発が加速している。
※4 ヘミセルロース:
水に不溶で薄いアルカリ溶液に溶ける植物細胞壁由来の炭水化物のこと。代表的なものとしてはマンナン、1,3-1,4-β-D-グルカン、キシラン、キシログルカンなどがあり、セルロースはβ-グルコースのみから構成されるのに対して、ヘミセルロースは複数の単糖によって構成されたヘテロ多糖であるという特徴がある。一般的にリグノセルロースに含まれる主要なヘミセルロースはキシランであり、β- D-キシロース分子が直鎖状に連なった主鎖がアラビノースやグルクロン酸側鎖等によって修飾されている。こうしたヘテロ多糖としての特性を利用して、さまざまな用途の糖源としての産業利用が模索されている。
※5 リグニン:
木材に含まれるフェノール性ポリマーであり、セルロースやヘミセルロースと化学結合することで、リグノセルロースに高い物理的・化学的強度を与えていると考えられている。紙パルプ製造プロセス、あるいはバイオエタノール製造プロセスの副生成物として工業的に得られてきたが、近年ではリグニン自体の高度利用開発が進んでおり、例えば、香料(バニリン)や接着剤、樹脂の原材料として利用されている。
※6 木質細胞:
植物細胞はすべて細胞壁をもっているが、植物個体の中でも特定の細胞のみが、細胞成長の完了後に新たにリグノセルロースを主体とした厚い細胞壁(二次細胞壁)を形成する。こうした細胞のことを木質細胞と呼び、木質バイオマスの生産の場となっている。主要な木質細胞としては、木部に存在する道管細胞と繊維細胞があり、道管細胞は水や無機塩類を運ぶ通水細胞として、繊維細胞は体を支える支持細胞として、それぞれ機能している。また、道管細胞の特徴として、二次細胞壁がらせん状やピット状といったパターンをもって蓄積する点が挙げられる。
※7 baculites 変異体:
今回の研究で新たに単離したシロイヌナズナ変異体であり、野生型で見られる道管細胞のらせん状の二次細胞壁蓄積が起こらなくなった変異体として見出された。この表現型から、らせん状に巻くはずの殻が巻かなくなり直線状の殻となった異常巻アンモナイトBaculites sp.から名前を取って、baculites と名付けられた。baculites 変異体では二次細胞壁のセルロースを合成する酵素遺伝子CESA7 内に1塩基の置換(突然変異)が起こっており、そのためにセルロース合成酵素活性が失われた結果、二次細胞壁でのセルロースの生合成が起こらなくなっている。
※8 微小管:
細胞内に見られる直径約 25 nm の繊維状の構造であり、チューブリンと呼ばれるタンパク質から構成される、細胞骨格の一種である。微小管は細胞の形態を維持・制御する他、植物細胞においては細胞壁生合成の際のセルロース合成装置の配置制御にも重要な役割を果たしている。
【解説図】
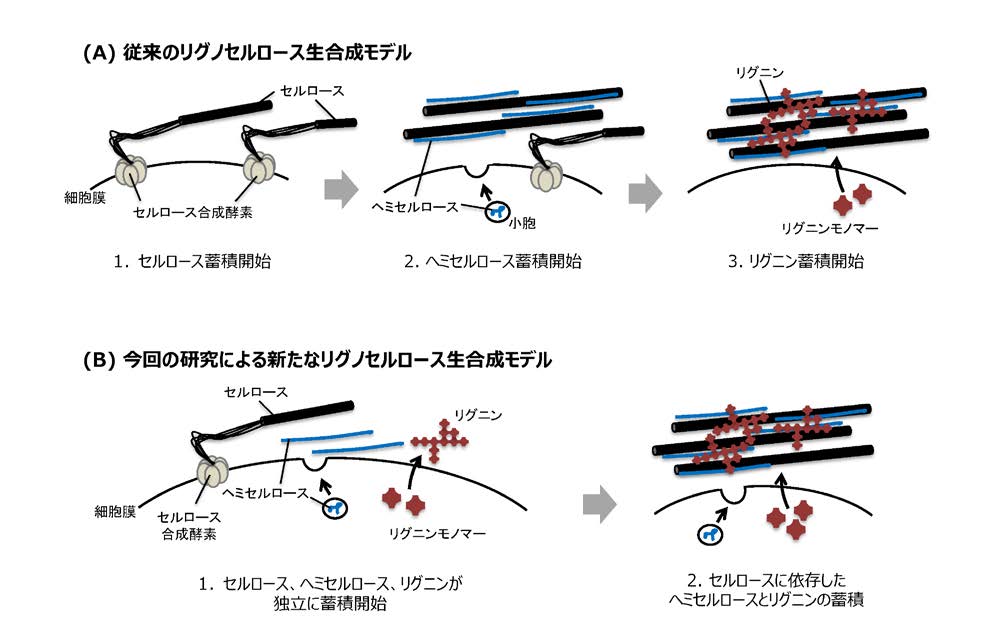
図1:リグノセルロース生合成モデル
(A) 従来から示されてきたリグノセルロース生合成モデル。これまでは、1.セルロース蓄積開始:セルロース合成酵素による細胞膜上でのセルロース合成が開始され、細胞壁中にセルロース蓄積が始まる、2.ヘミセルロース蓄積開始:ゴルジ体で合成されたヘミセルロースが細胞外へ小胞輸送され、細胞壁中に蓄積する、3.リグニン蓄積開始:細胞内で生合成されたリグニンモノマーが細胞外へ輸送され、細胞壁中で重合酵素によって重合する、と考えられており、2.と3.のステップはその前に起こるセルロース蓄積に依存すると考えられてきた。
(B)今回示すことに成功した新しいリグノセルロース生合成モデル。今回の成果によって、1.セルロース、ヘミセルロース、リグニンが独立に蓄積開始:木質細胞分化が始まると、それぞれが微小管に依存して蓄積を開始する、2.セルロースに依存したヘミセルロースとリグニンの蓄積:蓄積したセルロースに依存したヘミセルロースとリグニンのさらなる蓄積が起こる、という新たなモデルの提出に成功した。
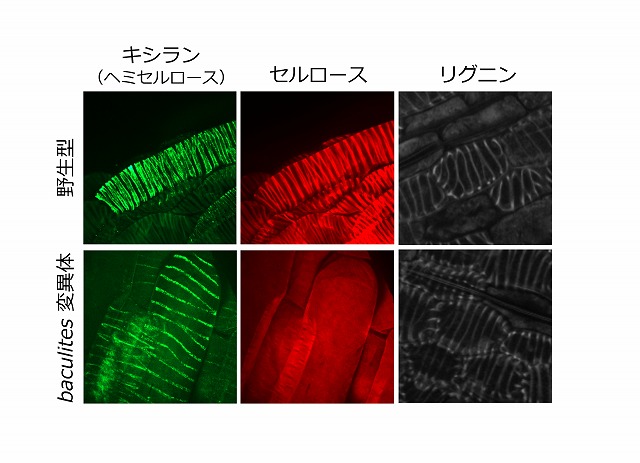
図2:セルロース生合成異常変異体baculitesではキシランとリグニンは蓄積している
道管細胞分化を誘導した野生型(上段)とbaculites(下段)の細胞について、キシラン(左:ヘミセルロース成分)、セルロース(中央)、リグニン(右)をそれぞれ可視化した。baculites変異体では、野生型で観察されるらせん状のセルロースシグナル(中央)が見られない一方で、キシラン(左)およびリグニン(右)のらせん状の存在を示すシグナルは検出された。
【植物代謝制御研究室】
研究室紹介ホームページ:http://bsw3.naist.jp/courses/courses104.html
研究室ホームページ:http://bsw3.naist.jp/demura/
(2018年11月06日掲載)
この記事をシェア ![]()