Research 研究成果の紹介
アブラナ目の生体防御を担う細胞の形成に気孔形成因子が転用されていた
~植物の特殊な機能を持つ細胞への進化の謎解明へ~
野菜の味の改変や二酸化炭素吸収能を高めた作物の開発に期待
アブラナ目の生体防御を担う細胞の形成に気孔形成因子が転用されていた
~植物の特殊な機能を持つ細胞への進化の謎解明へ~
野菜の味の改変や二酸化炭素吸収能を高めた作物の開発に期待
【概要】
奈良先端科学技術大学院大学(学長:塩﨑一裕)先端科学技術研究科 バイオサイエンス領域の白川一助教、山口暢俊准教授、伊藤寿朗教授、京都大学(総長:湊長博)大学院生命科学研究科 統合生命科学専攻の山岡尚平准教授、河内孝之教授らは、植物の葉の気孔を開閉する孔辺細胞と生体防御を担うミロシン細胞という2つの異なる機能に特殊化した細胞分化(注1)について、それぞれの細胞の遺伝子の発現に共通して関与する新規の転写因子(注2)である WASABI MAKER(WSB)を同定することに成功しました。さらに、植物が進化の過程で形成してきた転写ネットワークの一部を転用(注3)することにより、同じWSB を使いながら、2種の特殊化した細胞を獲得するというネットワークの仕組みを明らかにしました。
植物が進化の過程で、動物細胞が持たない特殊化した細胞を獲得してきたことはよく知られていますが、その遺伝子発現の仕組みの変遷を突き止めるための分子メカニズムはほとんど知られていませんでした。研究グループは、植物のガス交換を担う気孔の「孔辺細胞」と、細胞が傷つくと生体防御のための辛み成分などの物質を生成する「ミロシン細胞」に着目しました。ミロシン細胞が作る辛み成分は植食性の昆虫や細菌に対して生体防御物質として働く一方で、マスタードやワサビの辛み成分としても知られています。本研究では、孔辺細胞とミロシン細胞の分化を担う転写ネットワークを同定し、それらがどのように進化したか解析しました。その結果、孔辺細胞を作るために植物が獲得した転写ネットワークの一部が転用されて、WSB を併用する形でミロシン細胞を作るための転写ネットワークが獲得されたことがわかりました。
今後、植物が多様に特殊化した細胞を作るようになった進化のメカニズムを明らかにするとともに、作物の辛み成分の量や質を改変する技術や二酸化炭素の取り込みを効果的に行う気孔を備えた植物 を開発する技術へと発展することが期待されます。
本研究成果は、Nature Plants 誌に 2025 年 2 月 24 日(月)午後 7 時(日本時間)に公開されます
(DOI:10.1038/s41477-025-01921-1)。
【背景と目的】
植物は、動物にはない特殊な細胞を獲得し、地球環境に適応してきました。しかし、植物が持つ特殊な細胞を作るための遺伝子や、限られた遺伝子数の中で、植物が多様で特殊な細胞を生み出してきた仕組みはほとんどわかっていません。研究グループでは植物の 2 つの特殊化した細胞である「孔辺細胞」と「ミロシン細胞」に着目し、研究を行ってきました。
孔辺細胞は、ほぼすべての陸上植物の葉の表面に分布し、ガス交換と水分の蒸散を行っています。一方で、ミロシン細胞はアブラナ目植物だけが作る特殊な細胞で生体防御のために働いています。具体的には、ミロシン細胞に含まれる酵素であるミロシナーゼが基質であるグルコシノレート(カラシ油配糖体)と反応すると、揮発性の忌避物質であるイソチオシアネートが作られます。グルコシノレートはミロシン細胞とは別の特殊化した細胞(S細胞)に蓄積されており、外敵により食害を受けて細胞が壊れるとはじめて反応性の高い忌避物質が作られるという巧妙な防御システムです。イソチオシアネートは植食性の昆虫や細菌に対して生体防御物質として働く一方で、マスタードやワサビの辛み成分としても知られています。ミロシナーゼとグルコシノレートの反応による生体防御系は別名「カラシ油爆弾」とも呼ばれています。
これまでの本研究グループと他のグループの研究から、生物の設計図であるDNAからそのコピーであるmRNAを作る際に働く「FAMA」と呼ばれる転写因子が孔辺細胞とミロシン細胞の分化に必要であることがわかっていました。FAMAは数百から数千の下流遺伝子(転写される側)の発現のスイッチのオンとオフを制御して、2つの細胞の分化を実行していると考えられていましたが、その実体は不明でした。また、FAMAが両方の細胞分化に必要であるものの、2 つの全く異なる機能を持つ細胞をどのように分化させているかも不明でした。
【研究の内容】
FAMAが転写因子であることから、網羅的遺伝子発現解析(注4)を用いてその下流で制御される遺伝子群を同定しました。その結果、FAMAの誘導後すみやかに約600個の遺伝子の発現が影響を受けることが明らかになりました。候補遺伝子をさらに絞り込むために、過去に報告されている網羅的遺伝子発現解析データを含む別の3つの遺伝子リストと組み合わせることで、FAMA の影響を強く受ける候補遺伝子を 1 つに絞り込み、WASABI MAKER 転写因子(WSB)を同定しました。クロマチン免疫沈降法(注5)とレポーターアッセイ(注6)により、FAMAがWSBの発現を開始する部位(プロモーター)に直接結合して、その発現制御を受けることが示されました。また、WSBからFAMAへの正のフィードバックを発見し、FAMAとWSBの機能が互いに依存していることを発見しました。次にwsbの機能欠損変異体をゲノム編集の手法の 1 つである CRISPR/Cas9(注7)を用いて作製し、ミロシン細胞の分化を調べたところ、wsb 変異体では fama 変異体と同様に、ミロシン細胞の形成が観察されませんでした
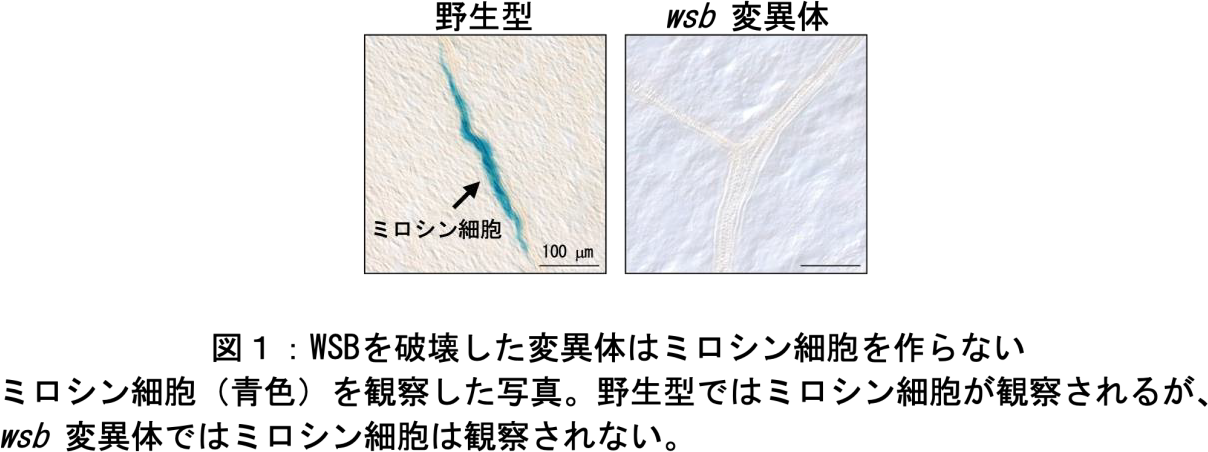
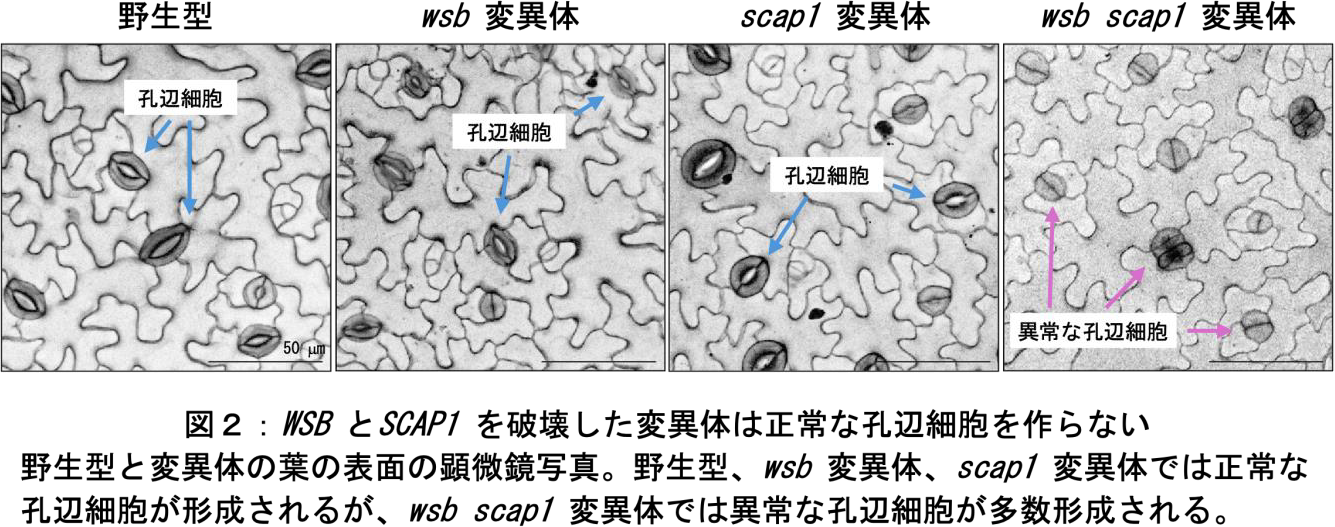
次に、wsb変異体の孔辺細胞の表現型を調べたところ、fama変異体のような異常は見られず、野生型(注8)と同様に分化していました。そこで、FAMAの下流標的因子を新たに探索し、Stomatal Carpenter 1(SCAP1)転写因子が孔辺細胞に特異的な FAMA の直接の標的因子であることを明らかにしました。scap1変異体もwsb変異体と同様に正常な孔辺細胞を形成しましたが、WSBとSCAP1の2つの転写因子の機能を破壊した植物(wsb scap1変異体)では、正常な孔辺細胞は野生型の3%程度しか観察されず、ほとんどが野生型の植物では見られない異常な孔辺細胞でした(図2)。以上の結果から、FAMAはWSBとSCAP1を介して孔辺細胞の分化を促進していることがわかりました。
また、植物の進化の過程でのWSB遺伝子の保存性を明らかにするために系統樹解析(注9)を行いました。その結果、WSBは被子植物に広く保存されていることがわかりました。このことから、WSBは被子植物の孔辺細胞分化に広く働いており、アブラナ目植物の出現時に、ミロシン細胞を作る機能も獲得したと考えられました。
そこで、アブラナ目植物のWSBがミロシン細胞を作るために新機能を獲得したと仮説を立てて実験を行ったところ、ミロシン細胞の分化において、WSB が SCAP1 を抑制することで、ミロシン細胞が「孔辺細胞らしさ」を獲得せずに済んでいることが明らかとなりました。WSB から SCAP1 への抑制制御は、孔辺細胞の形成システムでは働いておらず、ミロシン細胞分化においてのみ WSB が持つ新機能を発見しました。
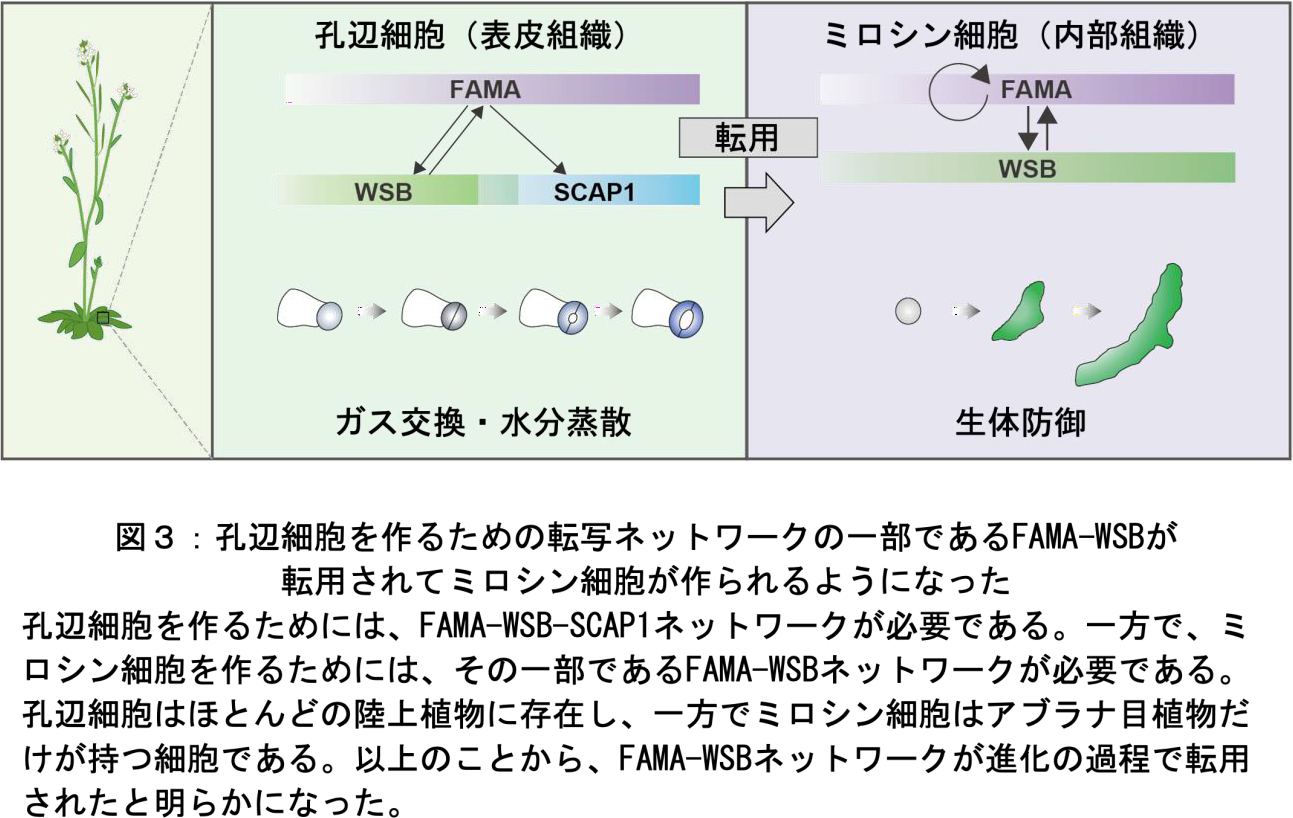
以上の結果から、FAMA-WSB-SCAP1 系が孔辺細胞分化で働いており、その転写ネットワークの縮小版である FAMA-WSB 系がミロシン細胞分化系に転用されたことが明らかとなりました。さらに、転用後WSB が新たな機能を獲得したことも示しました。植物が「転写因子ネットワークの転用」及び「WSB 転写因子に新機能を付与する」ことで、進化の過程で新規の特殊化した細胞を獲得したことが明らかになりました(図 3)。
【今後の展開】
今後は、FAMA と WSB に次いで稼働する下流標的遺伝子の同定と機能解析や、FAMA と WSB の発現部位を決める塩基配列(シス配列)の同定とその進化を明らかにすることで、植物が多様な特殊化した細胞を作るようになったメカニズムを明らかにする必要があります。また、FAMA やWSB のようなマスター因子(注 10)を野菜などにおいて改変することで、辛み成分の量を改変する技術や二酸化炭素の取り込みを効果的に行う植物を開発する技術へと発展することが期待されます。
【謝辞】
本研究は、日本学術振興会(JSPS)科学研究費補助金、特別研究員奨励費(12J05453)、新学術領域研究・植物の周期と変調(20H05416, 22H04723, 22H04726, 19H05670, 19H05675)、学術変革領域 B・天然物生物学(24H00881)、植物生殖改変(20H05780)、若手研究(18K14678)、基盤研究 C(19K06723)、基盤研究 B(22H02648, 21H02511, 18H02465, 23H02503)、基盤研究 A (JP20H00470, JP21K19266)、
学術変革領域A・不均一環境と植物(21H05665, 21H05663)、植物気候フィードバック(23H04968)、 学術変革領域 A・挑戦的両性花原理(22H05176)、科学技術振興機構(JST)さきがけ(JPMJPR22D3)、同 CREST(JPMJCR19S6)、セコム科学技術振興財団、武田科学振興財団、加藤記念バイオサイエンス振興財団、大隅基礎科学創生財団の支援を受けて行いました。
【用語解説】
注 1 細胞分化:もともと同じような性質を持つ細胞が、それぞれ異なる役割を持つ細胞に変化すること。
注 2 転写因子:遺伝子の転写(遺伝情報の RNA への複写)を調節するための分子のこと。これらの因子は DNA 上の特定の領域に結合し、遺伝子の活性化または抑制を制御し、生物の発達、成長、及び応答に影響を与える。
注 3 転用:特定の組織や細胞で発現していた遺伝子が、別の種では、もともとの特定の組織や細胞に加えて、別の組織や細胞で発現し、働くようになること。
注 4 網羅的遺伝子発現解析:生物の全ての遺伝子の発現パターンを同時に評価する技術。RNA シークエンシングなどの方法を使用して、特定の条件下でどの遺伝子がどれだけ発現しているかを定量的に測定する。
注 5 クロマチン免疫沈降法:タンパク質と DNA の相互作用を調べる生化学的手法の 1 つ。特定のタンパク質に特異的な抗体を使用して、そのタンパク質がクロマチン内で結合する特定の DNA 領域を検出する。
注 6 レポーターアッセイ:遺伝子が働いている組織や細胞を個体レベルで調べる方法。遺伝子が働く部位や、タイミングを明らかにすることができる。
注 7 CRISPR/Cas9:ゲノム編集法の 1 つ。DNA 配列を特定の方法で変更する技術の総称。CRISPR/Cas9 は主要な方法の 1 つで、疾患の治療や農作物の改良など多くの分野で応用される。
注 8 野生型:自然界に存在する標準的な遺伝子型や形質のことを指す。特に遺伝子変異がない、正常な状態の生物のこと。
注 9 系統樹解析:生物の進化的な関係を明らかにするために行う分析のこと。DNA やタンパク質の配列を比較し、それらがどのように共通の祖先から分岐してきたかを推定する。
注 10 マスター因子:細胞の分化や発生の際に、他の多くの遺伝子の発現を制御し、大きな影響を与える転写因子のこと。つまり、「細胞の運命を決める司令塔」。
【掲載論文】
タ イ ト ル :Co-option and neofunctionalization of stomatal executors for defense against herbivores in Brassicales
著 者 :Makoto Shirakawa1,2*, Tomoki Oguro1†, Shigeo S. Sugano3†, Shohei Yamaoka4†, Mayu Sagara1, Mai Tanida1, Kyoko Sunuma1, Takuya Iwami1, Tatsuyoshi Nakanishi1, Keita Horiuchi1, Kie Kumaishi5, Soma Yoshida6, Mutsumi Watanabe1, Takayuki Tohge1, Takamasa Suzuki7, Yasunori Ichihashi2,5, Atsushi Takemiya6, Nobutoshi Yamaguchi1, Takayuki Kohchi4, Toshiro Ito1*
†同等貢献、*責任著者
1:奈良先端科学技術大学院大学
2:科学技術振興機構(JST)さきがけ
3:産業技術総合研究所
4:京都大学
5:理化学研究所
6:山口大学
7:中部大学
掲 載 誌 :Nature Plants DOI:10.1038/s41477-025-01921-1
【花発生分子遺伝学研究室】
研究室紹介ページ:https://bsw3.naist.jp/courses/courses112.html
研究室ホームページ:https://bsw3.naist.jp/ito/
(2025年02月26日掲載)
この記事をシェア ![]()