Research 研究成果の紹介
一酸化窒素(毒性化合物)から酵母を保護する機構を解明
~抗真菌薬の開発や産業酵母の育種に期待~
一酸化窒素(毒性化合物)から酵母を保護する機構を解明
~抗真菌薬の開発や産業酵母の育種に期待~
【概要】
奈良先端科学技術大学院大学(学長:塩﨑一裕)先端科学技術研究科バイオサイエンス領域ストレス微生物科学研究室の那須野亮助教、高木博史教授のグループは、毒性の一酸化窒素(NO)による攻撃(NOストレス)から酵母を防御する新たなメカニズムを明らかにしました。NOストレスは、病原菌の感染過程や有用物質の発酵生産にも深く関わっているため、本研究による新たなNO耐性機構の解明は、新たな抗真菌薬の開発や発酵力を強化した産業用酵母の育種に貢献することが期待されます。
酵母Saccharomyces cerevisiaeは、発酵・醸造食品、バイオエタノールなどを生産する有用微生物であり、高等生物や病原性真菌などのモデル生物としても研究されています。発酵環境下では、酵母がNOストレスを受ける場合があるため、NOに対する耐性機構を理解することは発酵産業において極めて重要です。一方、病原性真菌が感染する際には、宿主の免疫細胞が合成するNOが病原性真菌を攻撃するため、病原性真菌のNOに対する耐性機構は新たな抗真菌薬の標的として注目されています。NOは生体分子であるタンパク質を化学的に修飾することで様々な機能を発揮しますが、NOストレスに対する酵母の挙動についてはあまり分かっていませんでした。
本研究では、ブドウ糖をエネルギーに変える解糖系の「フルクトース二リン酸アルドラーゼFba1」と言う酵素が、NO依存的な化学修飾であるS-グルタチオン化(GSH化)されることで、その活性が阻害されることを見出しました。さらに、Fba1の阻害により、解糖系のバイパス経路であるペントースリン酸回路(PPP)が活性化され、その結果、NOの解毒に重要なNADPHがPPPにより多量に合成されることで、酵母のNO耐性が亢進することを明らかにしました。
本研究で明らかにしたGSH化修飾を介した代謝制御によるNO耐性機構はこれまでに報告がなく、酵母におけるNO応答・耐性機構のさらなる理解に大きく貢献します。また、NO耐性に関する知見に基づいた抗真菌薬の開発や高い発酵力を有する酵母の育種につながることが期待されます。
本研究の成果は、オープンアクセスの国際学術誌「Free Radical Biology and Medicine誌」のオンライン版に、2022年10月20日(木)付で掲載されました。
那須野亮助教のコメント

一酸化窒素(NO)は、哺乳類の血管内皮細胞を弛緩させる因子として発見されて以降、様々な生物において研究が行われてきました。しかし、酵母におけるNOの機能やその分子機構については研究例・知見が少なく、理解が進んでいませんでした。本研究では、GSH化という翻訳後修飾を切り口に酵母のNO耐性機構を解明することにより、酵母におけるNOの機能をより深く理解するための重要な知見を得ることができました。一方、本研究の成果は、酵母のNO耐性機構を解明することで、発酵生産の人為的制御や新しい抗真菌薬の開発にも貢献することが期待できます。
本研究は、バイオサイエンス領域ストレス微生物科学研究室において実施されました。高木博史教授には、本研究に集中して取り組むための環境を物心両面からサポートしていただきました。また、酵母を基礎・応用の両面から研究することの楽しさと重要性を熱心にご指導いただいております。一方、当研究室の修了生で今年3月に博士学位を取得した示野誠也君は、粘り強く丁寧な実験や深い考察・議論により、本研究成果の創出に大きく貢献してくれました。また、本領域の技術職員である倉田理恵さんには、プロテオーム解析に関する相談から解析まで大変お世話になりました。皆様方には、この場を借りまして心より感謝申し上げます。
【解説】
[研究背景]
哺乳類で発見された一酸化窒素(NO)は、シグナル分子として血圧の調節や神経伝達など様々な生命現象に関与しています。一方、過剰な量のNOは細胞死や細胞機能障害(NOストレス状態)を引き起こすため、多くの生物にはNOに対する防御機構が備わっています。また、NOは様々な分子と化学反応を起こし、特にタンパク質に翻訳後修飾と呼ばれる多様な化学修飾を引き起こします。
酵母Saccharomyces cerevisiaeは、発酵・醸造食品やバイオエタノールなどの発酵生産に用いられる重要な微生物です。発酵環境下では、培地成分として用いられる廃糖蜜に含まれる亜硝酸塩からNOが合成され、酵母の発酵力は制限されます(図1A)。一方、酵母は病原性真菌のモデル生物としても研究されています。病原性真菌が動物に感染する場合、宿主である動物の免疫細胞がNOを合成し、その反応性により病原性真菌を攻撃・殺菌することで、感染を防御しています(図1B)。以上のように、酵母のNO耐性を解明することは、酵母の発酵力の制御や病原性真菌の感染戦略の理解につながります。
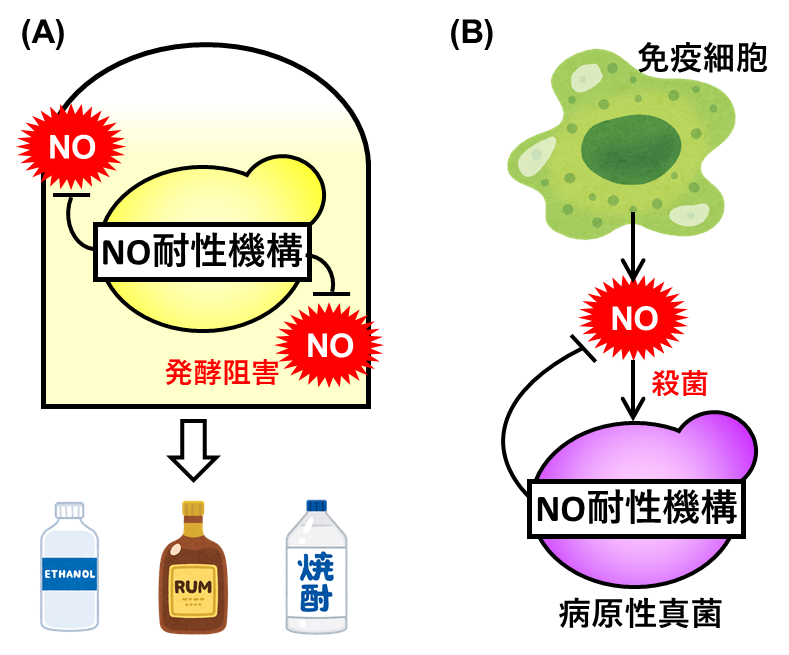
図1 酵母のNO耐性と発酵・感染の関連性
(A)酵母は発酵環境下において、炭素源として用いられる廃糖蜜などに含有する亜硝酸塩由来のNOに曝され、発酵力が制限される。そのため、酵母の発酵力の制御や効率化のためには、酵母のNO耐性の理解が重要である。(B)酵母の近縁種である病原性真菌は、感染時に宿主の免疫細胞が合成するNOにより攻撃・殺菌される。一方、病原菌はNOによる攻撃を回避するためのNO耐性を有しており、感染戦略として機能する。したがって、病原性真菌のNO耐性機構は新たな抗真菌薬の標的となる。
酵母のNOに対する耐性・応答機構については、NO分解酵素などが報告されているものの、あまり研究は進んでいません。特に、NOによるタンパク質の翻訳後修飾についてはほとんど知見がありませんでした。そこで、本研究ではNO依存的な翻訳後修飾を受けるタンパク質を明らかにし、NO耐性との関連について解明することを目的としました。
[研究結果]
タンパク質の翻訳後修飾を網羅的に解析するプロテオーム解析を行ったところ、ブドウ糖をエネルギーに変換する解糖系の酵素「フルクトース二リン酸アルドラーゼFba1」のシステイン残基が、NOストレスに応答して何らかの翻訳後修飾を受けることを見出しました。さらに、翻訳後修飾を特異的に検出するウェスタンブロット法により解析を行ったところ、Fba1のシステイン残基がS-グルタチオン化(GSH化)修飾を受けることを見出しました。続いて、システイン残基をGSH化が起こらないセリン残基に置換したFba1を発現する酵母を用いて解析を行った結果、NOストレスによりFba1の112番目のシステイン残基(Cys112)がGSH化を受けることを明らかにしました(図2)。
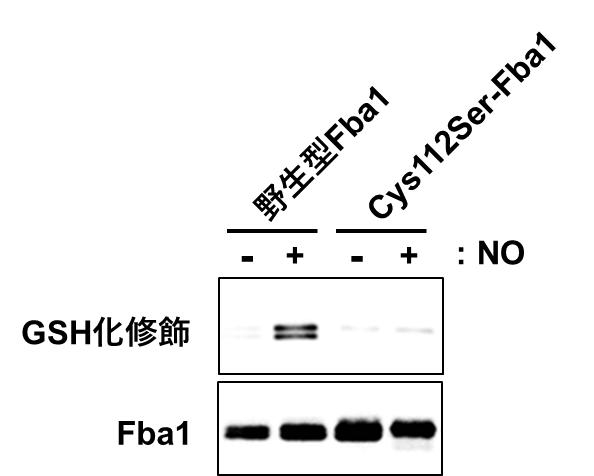
図2 NOストレスによるFba1のGSH化
NOストレスで処理した酵母の抽出液からFba1を単離し、GSH化を特異的に検出するウェスタンブロット解析によって、Fba1のGSH化修飾を解析した。野生型Fba1はNOストレスによりGSH化されるが、Cys112Ser-Fba1はGSH化されないことから、NO処理によってFba1のCys112残基が特異的にGSH化されることが明らかになった。
大腸菌を用いて組換えタンパク質として発現・精製したFba1を用いて酵素活性を測定したところ、化学的にGSH化修飾を導入することによりFba1の酵素活性が抑制されることを見出しました(図3)。GSH化修飾は還元酵素グルタレドキシンにより脱修飾されることが知られています。酵母が持つ4種類の細胞質型グルタレドキシンを用いた解析から、グルタレドキシンの一種Grx1がFba1のGSH化と酵素活性の抑制を解除することを明らかにしました。
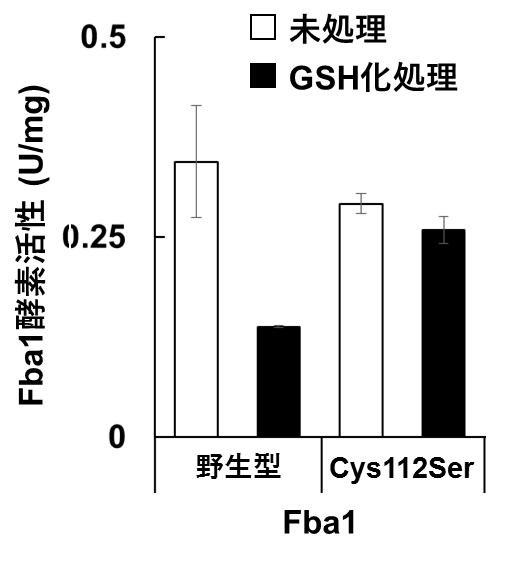
図3 GSH化によるFba1酵素活性の抑制
大腸菌を用いた組換えタンパク質としてFba1を精製し、酵素活性を測定した。化学的処理によりGSH化修飾を誘導したところ、野生型Fba1の酵素活性は半分以下に低下した。一方、GSH化が起こらないCys112Ser-Fba1の活性はGSH化処理により低下しなかった。
Fba1は解糖系に必須の酵素であるため、Fba1が抑制された条件では、ブドウ糖は別の経路で代謝されると予想されました。そこで、Fba1の基質であるフルクトース-1,6-二リン酸(FBP)、解糖系のバイパス経路であるペントースリン酸回路(PPP)の代謝物である6-ホスホグルコン酸(6PG)の細胞内含量を測定したところ、NOストレスに応答してこれらの代謝物量が増加しました(図4A, B)。一方、Cys112をセリン残基に置換してGSH化が起こらないようにしたFba1(Cys112Ser-Fba1)を発現する酵母では、これらの代謝物の細胞内含量はNOストレスによって変化しませんでした。これらのことから、NOストレス処理によってFba1がGSH化を受けることにより、代謝の流れが解糖系からPPPへと変化することが明らかになりました。PPPで合成されるNADPHは既知のNO耐性機構であるNO分解酵素の活性に必要な化合物です。酵母細胞内のNADPH量はNO処理によって増加しましたが、Cys112Ser-Fba1を発現する酵母ではNADPHは増加しませんでした(図4C)。さらに、Cys112Ser-Fba1を発現する酵母は野生型の酵母よりもNOストレスに対して感受性を示しました。
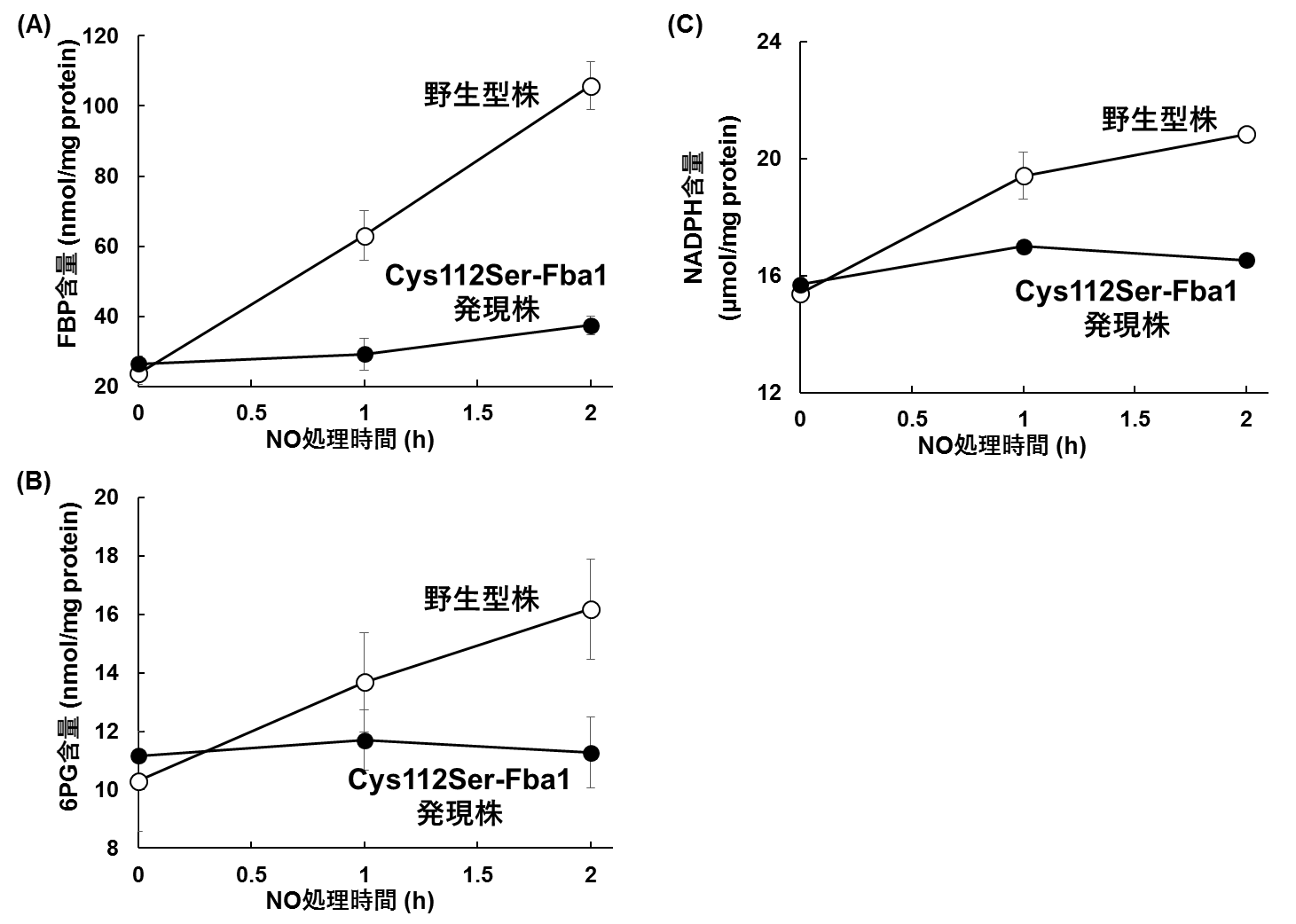
図4 NOに応答した代謝物量の変化
酵母の野生型株、およびGSH化が起こらないCys112Ser-Fba1を発現する株をNOストレスで処理し、細胞内の(A)FBP含量、(B)6PG含量、(C)NADPH含量を測定した。各代謝物とも、野生型株においてはNO処理に応答して増加したが、GSH化が起こらないCys112Ser-Fba発現株では代謝物量の増加は見られなかった。
以上の結果から、酵母がNOストレスに曝された際、Fba1のCys112がGSH化修飾を受けることでFba1の酵素活性が抑制され、代謝が解糖系からPPPへと変化した結果、NADPH合成が亢進したことにより、酵母のNO耐性が強化されるということが解明されました(図5)。
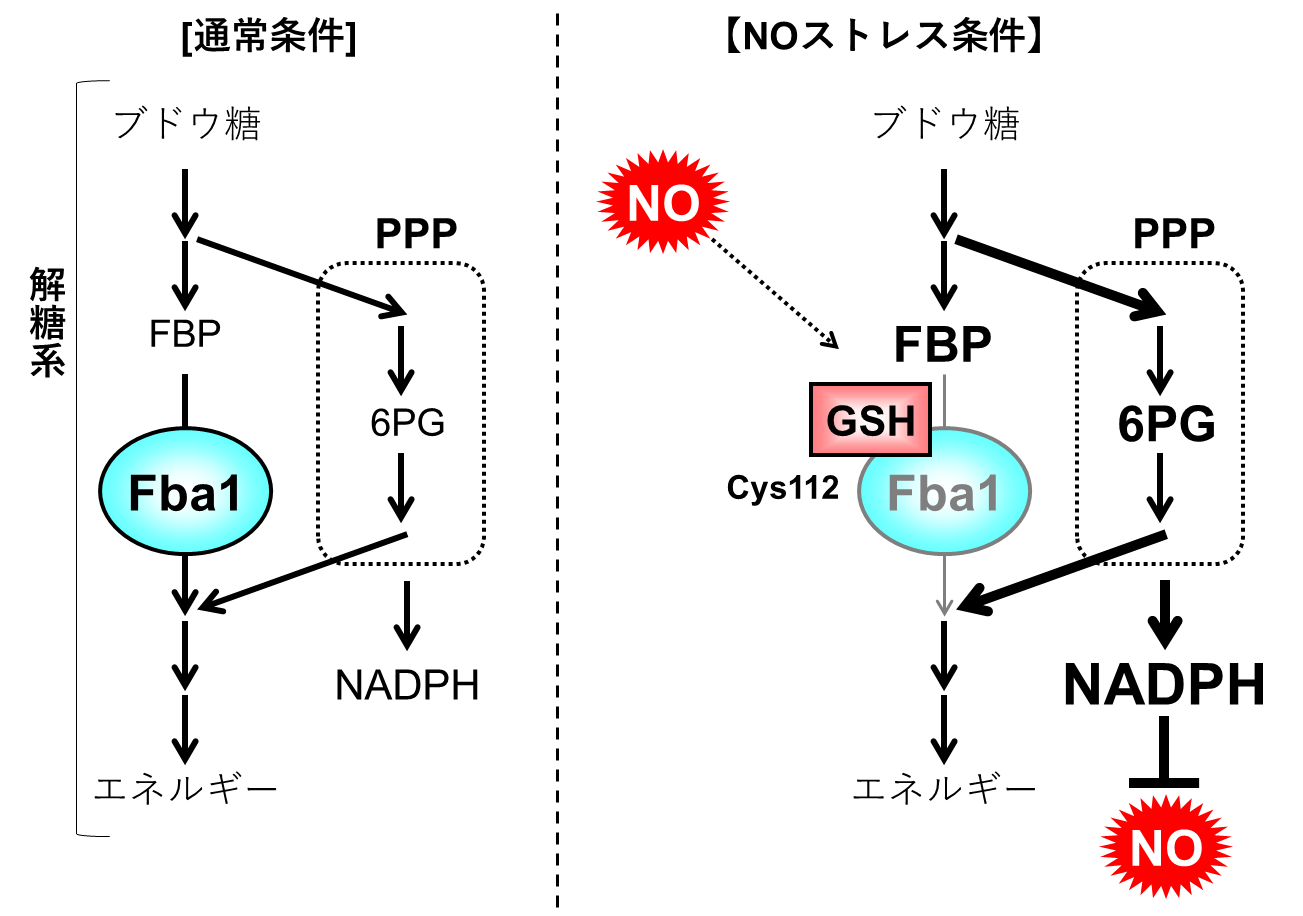
図5 Fba1のGSH化による代謝変化を介したNO耐性機構
酵母がNOストレスに曝されると、Fba1のCys112残基がGSH化修飾を受けることにより、Fba1の酵素活性が抑制される。その結果、解糖系からPPPへと代謝の流れが変化し、PPPにおけるNADPHが亢進することにより、酵母がNO耐性を獲得する。
[研究意義]
酵母におけるNO耐性やNO応答のメカニズムには不明な点が多く、特に翻訳後修飾の生理的意義や分子機構はほとんど分かっていませんでした。今回の研究で見出したGSH化修飾による代謝制御を介したNO耐性機構はこれまでにない新たなメカニズムであり、酵母におけるNOの機能のさらなる理解に大きく貢献します。
発酵生産における炭素源として用いられる廃糖蜜には高濃度の亜硝酸塩が含有されており、NOストレスにより発酵が阻害されると考えられています(図1A)。今回の研究で見出したFba1依存的なNO耐性機構を人為的に改変・強化することにより、NOストレス下でも発酵力が低下しない有用酵母の育種が可能になると期待されます。
病原性真菌のNO耐性機構は、免疫細胞が合成するNOによる攻撃を回避するための感染戦略であるため、抗真菌薬の標的として極めて有望です(図1B)。また、抗真菌薬開発においては、副作用の低減のため、薬剤標的が病原性真菌にのみ存在することが重要です。フルクトース二リン酸アルドラーゼは、哺乳類や高等植物に保存されるI型と、酵母や真菌、細菌などに保存されるII型に分類され、それぞれ特有の構造や反応機構を有しています。Fba1のCys112もII型のフルクトース二リン酸アルドラーゼにのみ保存されるため、今回の研究で見出したFba1依存的なNO耐性機構は微生物に特有のメカニズムと考えられます。そのため、Fba1のGSH化を特異的に阻害する化合物は、新たな抗真菌薬の有力な候補となることが考えられます。
【掲載論文】
タイトル:S-glutathionylation of fructose-1,6-bisphosphate aldolase confers nitrosative stress tolerance on yeast cells via a metabolic switch
(和訳:フルクトース二リン酸アルドラーゼのS-グルタチオン化は代謝変換を介して酵母にNO耐性を付与する)
著者:Seiya Shino, Ryo Nasuno*, Hiroshi Takagi*
* Corresponding author
掲載誌:Free Radical Biology and Medicine
DOI: 10.1016/j.freeradbiomed.2022.10.302
[用語解説]
・一酸化窒素(NO)
NOは哺乳類で血管拡張因子として見出されたガス状の活性分子であり、様々な生命現象に関与する。一方、過剰量のNOは細胞毒性を有するため、生物は様々なNO耐性機構を有する。NOはアルギニンや硝酸塩、亜硝酸塩から酵素的に合成されるほか、亜硝酸塩から非酵素的な化学反応により生成する。例えば、動物の免疫細胞はアルギニンからNOを合成し、病原菌を攻撃することにより感染を防ぐ。また、発酵生産に用いられる培地成分に混入する亜硝酸塩はNOの発生源となり、発酵生産を抑制すると考えられている。
・酵母
酵母は、遺伝解析やゲノム解析による基礎的知見が蓄積され、遺伝子操作技術も確立されているため、ヒトなどの高等生物や病原性真菌のモデル生物として研究されている。一方、パン類や酒類などの発酵・醸造食品、バイオエタノールなどの生産に用いられる重要な産業用微生物である。
・翻訳後修飾
DNAにコードされる遺伝情報はmRNAへと転写され、それをもとにアミノ酸が連なったタンパク質が合成される。タンパク質の合成(翻訳)の後にタンパク質のアミノ酸残基が化学的に修飾を受ける現象は翻訳後修飾と呼ばれる。様々な翻訳後修飾が知られており、タンパク質の構造や機能を制御することで多様な生命現象に関わる。
・S-グルタチオン化
システイン残基における翻訳後修飾の一種。抗酸化機能を持つトリペプチドであるグルタチオンとシステイン残基がジスルフィド結合を形成した修飾。還元剤やグルタレドキシンなどの還元酵素により、未修飾のタンパク質へと脱修飾される。
・プロテオーム解析
タンパク質の存在量や翻訳後修飾を網羅的に解析する手法。タンパク質を酵素分解したものを液体クロマトグラフィーと質量分析に供し、得られたシグナルをデータベースの情報と照合することで、タンパク質の同定・定量、翻訳後修飾の解析などを行うことができる。
【ストレス微生物科学研究室】
研究室紹介ホームページ:http://bsw3.naist.jp/courses/courses305.html
研究室ホームページ:http://bsw3.naist.jp/takagi/
(2022年11月10日掲載)
この記事をシェア ![]()