Research 研究成果の紹介
根端の細胞が自ら剥がれ落ちるしくみを解明
 バイオサイエンス研究科・植物発生シグナル研究室の中島敬二教授らの研究グループは、理化学研究所との共同研究により、根の先端を覆う根冠組織から、生きた細胞が規則的に剥がれ落ちる仕組みを明らかにしました。根冠細胞の剥離は、根の成長のみならず、土壌環境の調節にも重要な役割を担っており、その仕組みの解明は、植物の成長力の改良や土壌環境の改善にもつながる研究成果です。この成果を掲載した論文は、11月1日(火)発行のDevelopmentに掲載されました。
バイオサイエンス研究科・植物発生シグナル研究室の中島敬二教授らの研究グループは、理化学研究所との共同研究により、根の先端を覆う根冠組織から、生きた細胞が規則的に剥がれ落ちる仕組みを明らかにしました。根冠細胞の剥離は、根の成長のみならず、土壌環境の調節にも重要な役割を担っており、その仕組みの解明は、植物の成長力の改良や土壌環境の改善にもつながる研究成果です。この成果を掲載した論文は、11月1日(火)発行のDevelopmentに掲載されました。
中島敬二教授による研究成果の解説
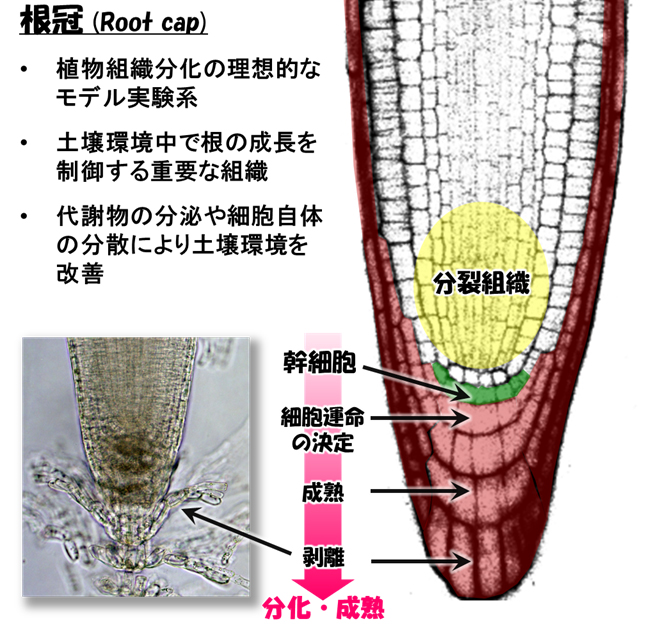
図1
根の先端には根冠(こんかん)と呼ばれるキャップ型の組織がはまっています。根冠はあまり目立たない組織ですが、根の成長に必須の様々な役割を担っています。例えば、根端の分裂組織は、根冠に包まれることで土壌との接触から守られています(図1)。また根冠は様々な物質を分泌して根の成長に適した土壌環境を作り出しています。根冠の中央部は重力感受細胞として働き、根の伸びる方向を調節しています。
根冠組織は植物種によって数層から十数層の細胞層からできていますが、その作られ方には、他の組織には見られないユニークな特徴があります。それは、根冠の内側で新しい細胞が作られ、外側の細胞から順に剥離してゆくことです(図1)。このように細胞が常に入れ替わることで、根冠という根のフロントラインには常に生命力の高い細胞が配置されます。このような細胞のターンオーバーはヒトの皮膚でも見られますが、根冠が皮膚と異なるのは、外側の細胞が一定の間隔で、自発的に、しかも生きたまま剥離することです。根冠組織の外側の細胞だけが、どうして規則正しく剥がれてゆくのか、その仕組みはこれまで分かっていませんでした。
私たちの研究室では、モデル植物のシロイヌナズナにおいて根冠細胞の分化を調節するSOMBRERO (SMB) 転写因子の機能を調べていました。その過程で、SMBが別の2つの転写因子BEARSKIN1 (BRN1)とBRN2 の発現を促進することを発見しました。これらの3つの転写因子は、根冠細胞の分化や成熟を制御することが報告されていましたが、その使い分けや、標的遺伝子は分かっていませんでした。
まず私たちは、SMBが根冠全体で発現しているのに対し、BRN1とBRN2は根冠の外側の1-2層のみで発現していることを発見しました。また、SMBを過剰発現させると、BRN1とBRN2の発現領域も広がりましたが、興味深いことに、SMBを根全体で過剰発現させても、BRN1とBRN2の発現は、根の外側に露出した表皮細胞にしか広がりませんでした。このことは、根の細胞が「個体の表面にある」ことを何らかの方法で感知し、そのような「位置情報」に応じてBRN1とBRN2の発現を活性化していることを示しています。
次にこれら3つの転写因子が調節している標的遺伝子を探索し、60個の候補遺伝子を見つけました。これらの中には植物の細胞壁に作用する酵素や、細胞外への分泌に働くタンパク質をコードする遺伝子が多く含まれており、根冠細胞が担う機能とよく一致していました。実際にこれらの遺伝子の少なくともいくつかは、根冠の外側の細胞で特異的に発現していました。
当時、大学院博士後期課程に在籍していた神谷雅子さんは、細胞壁の主成分の1つであるペクチンを分解するポリガラクツロナーゼをコードする遺伝子に注目し、この遺伝子をRCPG (ROOT CAP POLYGALACTURONASE)と名付けて詳細に解析しました。ペクチンは細胞壁を構成する主要な多糖成分の1つであり、セルロース繊維の間を埋めるゲル状の物質です。ポリガラクツロナーゼはペクチンの主鎖を構成するポリマーを加水分解して切断する酵素ですから、RCPGが根冠細胞の剥離に重要な機能を果たしているのではないかと推測したのです。
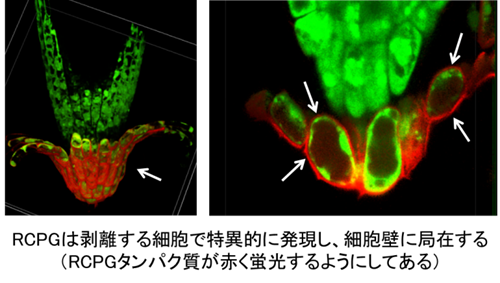
図2
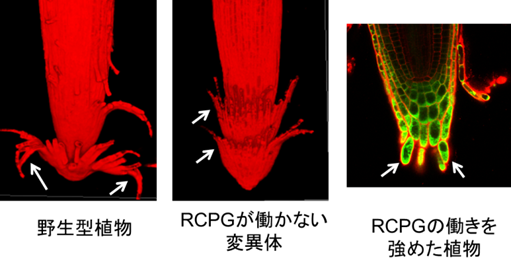
図3
まず神谷さんは、RCPGタンパク質が根冠の最外層の細胞で急激に発現し、それらの細胞の細胞表面へ運ばれることを突き止めました(図2)。一方で、内側の根冠細胞や根の他の組織では、RCPGタンパク質はほとんど発現していませんでした。この発現パターンはBRN転写因子と類似していたため、両者の関係を調べたところ、BRN1とBRN2を同時に破壊した変異体では、RCPGの発現が極端に低下していました。また、少なくともBRN1転写因子は、RCPGプロモーターに直接結合して転写を活性化していることを、理化学研究所の金博士らとの共同研究で明らかにしました(ちなみに金博士も本研究科の修了生です)。
次に神谷さんは、RCPG遺伝子の変異体(rcpg変異体)を単離し、根冠細胞の様子を野生型と比較しました。その結果、野生型植物では、根冠の最外層が大きく反り返って剥がれていたのに対し、rcpg変異体では、根冠の最外層がカップ状の外形を維持し、根端にはまったままになっていました(図3)。反対にRCPGの過剰発現体では、根冠の最外層の細胞が層構造を保持せずに、バラバラと個別に剥がれ落ちているのが観察され、これによってRCPGが細胞の剥離を促進していることが確かめられました(図3)。
以上の実験結果から、次のような一連の機構が繰り返されることが推定されます(図4)。
(1) SMBを発現している根冠細胞が、根の表面にいることを感知する
(2) その情報をもとにBRN1とBRN2転写因子が根の最外層で発現する
(3) BRN1とBRN2がRCPG遺伝子の転写を活性化してペクチン分解酵素を作る
(4) 生成したペクチン分解酵素が細胞壁に運ばれてペクチンを分解する
(5) 細胞接着が緩んで最外層の細胞が生きたまま剥離する
(6) 外から2層目にあった細胞が最外層となって(1)のステップに戻る
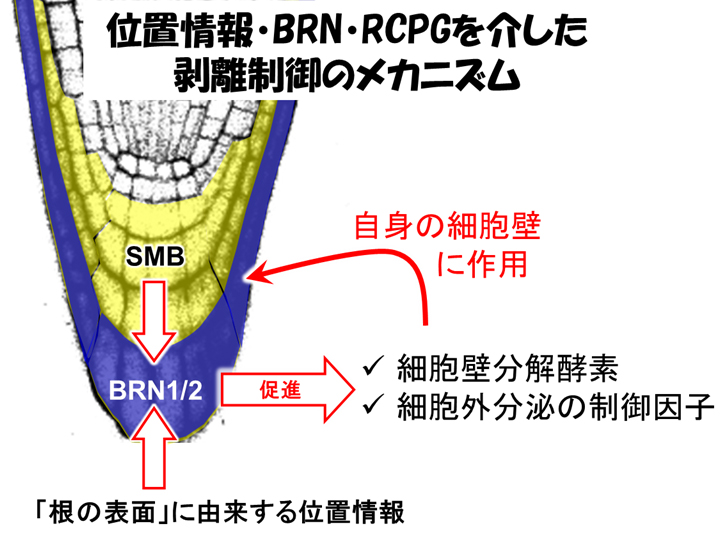
図4
こうした仕組みにより根冠細胞が生きたまま剥がれることで、細胞内にたまった様々な代謝産物が分解されることなく直接土壌中に散布されます。これらの作用により整えられた土壌環境の中を根が伸長してゆくことで、栄養分の効率的な吸収が促進されたり、病原菌の感染が抑えられることが想像されます。
この論文では、根冠の機能のうちでも特に細胞の剥離に焦点を絞りましたが、今回の研究では、他にも興味深い遺伝子がいくつか見つかっています。今後はこれらの働きを順次解明してゆくことで、根冠細胞の様々な機能が、どのように発揮されるかを明らかにできると期待しています。
【論文情報】
タイトル:
Control of root cap maturation and cell detachment by BEARSKIN transcription factors in Arabidopsis
Masako Kamiya, Shin-ya Higashio, Atsushi Isomoto, Jong-Myong Kim, Motoaki Seki, Shunsuke Miyashima, and Keiji Nakajima
Development 143: 4063-4072 (2016); http://dev.biologists.org/content/143/21/4063
【植物発生シグナル】
研究室紹介ページ:http://bsw3.naist.jp/courses/courses110.html
研究室ホームページ:http://bsw3.naist.jp/nakajima/
(2016年11月07日掲載)
この記事をシェア ![]()