Research 研究成果の紹介
オーキシンが発根を促進するメカニズムを解明 〜発根を調節する農薬の開発に期待〜
オーキシンが発根を促進するメカニズムを解明〜発根を調節する農薬の開発に期待〜
名古屋大学大学院生命農学研究科(研究科長:川北一人)の古谷将彦(ふるたにまさひこ)研究員は、このたび、奈良先端科学技術大学院大学(学長:小笠原直毅)バイオサイエンス研究科の田坂昌生(たさかまさお)教授、同研究科の井藤純(いとうじゅん)助教、神戸大学大学院理学研究科の深城英弘(ふかきひでひろ)教授、中国科学院との共同研究において、植物の発根を促進するホルモンであるオーキシンに係るその下流で働く遺伝子を活性化する基本的な仕組みを明らかにしました。
これまで、オーキシンと結合した受容体が、転写抑制因子(Aux/IAA)を分解することで、下流遺伝子の転写をONにすると考えられていました。転写に
は、転写装置(RNAポリメラーゼⅡ)の起動が必要なのですが、オーキシンによるAux/IAAの分解が転写装置を起動する仕組みは謎のままでした。
本研究では、転写因子と転写装置をつなぐ転写メディエーター複合体に着目し、オーキシンがAux/IAAを分解し転写メディエーター複合体のかたちを変えることで、転写装置に動力を伝達することを発見しました。
本研究の成果により、今後、オーキシンと同じように転写メディエーター複合体のかたちを変えることができる化学物質を発根促進剤として開発することで、希少・有用植物の挿木・挿苗による量産化が期待されます。 この研究成果は、平成28年5月23日付(米国東部時間15時)米国科学雑誌「米国科学アカデミー紀要」オンライン版に掲載されました。
古谷将彦研究員のコメント
本研究は、私がバイオサイエンス研究科に所属していた期間に(今年 度3月末まで)当研究科の田坂昌生教授と井藤純助教との共同で行われました。7年前の修了生曽野貴子さんが同定したMAB2遺伝子(転写メディエーター複合体の構成因子MED13をコード)を足がかりに、筆頭著者の井藤純助教が中心となってオーキシンシグナル伝達における転写メディエーター複合体の機能を明らかにしました。これまでにオーキシン受容体TIR1が転写抑制因子Aux/IAAをオーキシン依存的に分解することで下流遺伝子の転写をOFFからONにスイッチすることが分かっていましたが、オーキシンが転写装置(RNAポリメラーゼ)をどのように制御するかについては明らかにされていませんでした。本研究では、転写因子と転写装置を仲介する転写メディエーター複合体の構成要素がオーキシンによって変化することで、下流遺伝子の転写のON・OFFが制御されることを実験的に証明しました。今後は、オーキシンと同じように転写メディエーター複合体の構成要素を制御する化学物質を探索し、新たな農薬の開発を目指します。
度3月末まで)当研究科の田坂昌生教授と井藤純助教との共同で行われました。7年前の修了生曽野貴子さんが同定したMAB2遺伝子(転写メディエーター複合体の構成因子MED13をコード)を足がかりに、筆頭著者の井藤純助教が中心となってオーキシンシグナル伝達における転写メディエーター複合体の機能を明らかにしました。これまでにオーキシン受容体TIR1が転写抑制因子Aux/IAAをオーキシン依存的に分解することで下流遺伝子の転写をOFFからONにスイッチすることが分かっていましたが、オーキシンが転写装置(RNAポリメラーゼ)をどのように制御するかについては明らかにされていませんでした。本研究では、転写因子と転写装置を仲介する転写メディエーター複合体の構成要素がオーキシンによって変化することで、下流遺伝子の転写のON・OFFが制御されることを実験的に証明しました。今後は、オーキシンと同じように転写メディエーター複合体の構成要素を制御する化学物質を探索し、新たな農薬の開発を目指します。
【ポイント】
●オーキシンが転写メディエーター複合体のかたちを変えることを発見。
●下流遺伝子のプロモーター上で転写メディエーター複合体のかたちが変化することに伴い、下流遺伝子の転写も変化することを確認。
●オーキシンが下流遺伝子の転写を調節する機構の全容解明に近づく。
【研究背景と内容】
植物の根は、図1のように内部から側方に側根を形成し、植物ホルモ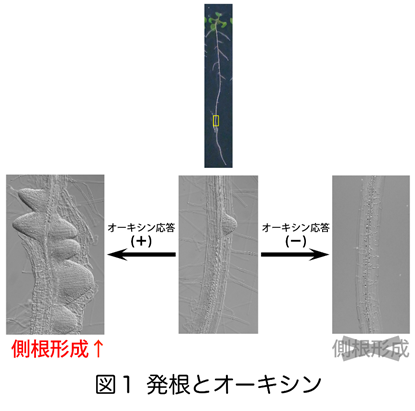 ンであるオーキシンがその形成を促進します。オーキシンに対する応答性が高くなるとたくさんの側根が生え、逆に低くなると側根が生えません(図1)。そのオーキシンの応答性を決定しているのは、たくさんの下流遺伝子の転写を調節するAUXIN RESPONSE FACTOR(ARF)と呼ばれる転写因子とAUXIN/INDOLE 3-ACETIC ACID(Aux/IAA)という転写抑制因子です。オーキシンが存在しない場合は、Aux/IAAがARFと結合しARFの転写因子としての役割を阻害します。
ンであるオーキシンがその形成を促進します。オーキシンに対する応答性が高くなるとたくさんの側根が生え、逆に低くなると側根が生えません(図1)。そのオーキシンの応答性を決定しているのは、たくさんの下流遺伝子の転写を調節するAUXIN RESPONSE FACTOR(ARF)と呼ばれる転写因子とAUXIN/INDOLE 3-ACETIC ACID(Aux/IAA)という転写抑制因子です。オーキシンが存在しない場合は、Aux/IAAがARFと結合しARFの転写因子としての役割を阻害します。
一方、オーキシンが存在する場合は、オーキシンと結合した受容体TIR1 (TRANSPORT INHIBITOR RESPONSE 1)がAux/IAAを分解し、ARFが転写活性化因子としての能力を発揮すると考えられてきました。オーキシンが側根を作る際にはAux/IAA14/SOLITARY-ROOT(SLR)が分解され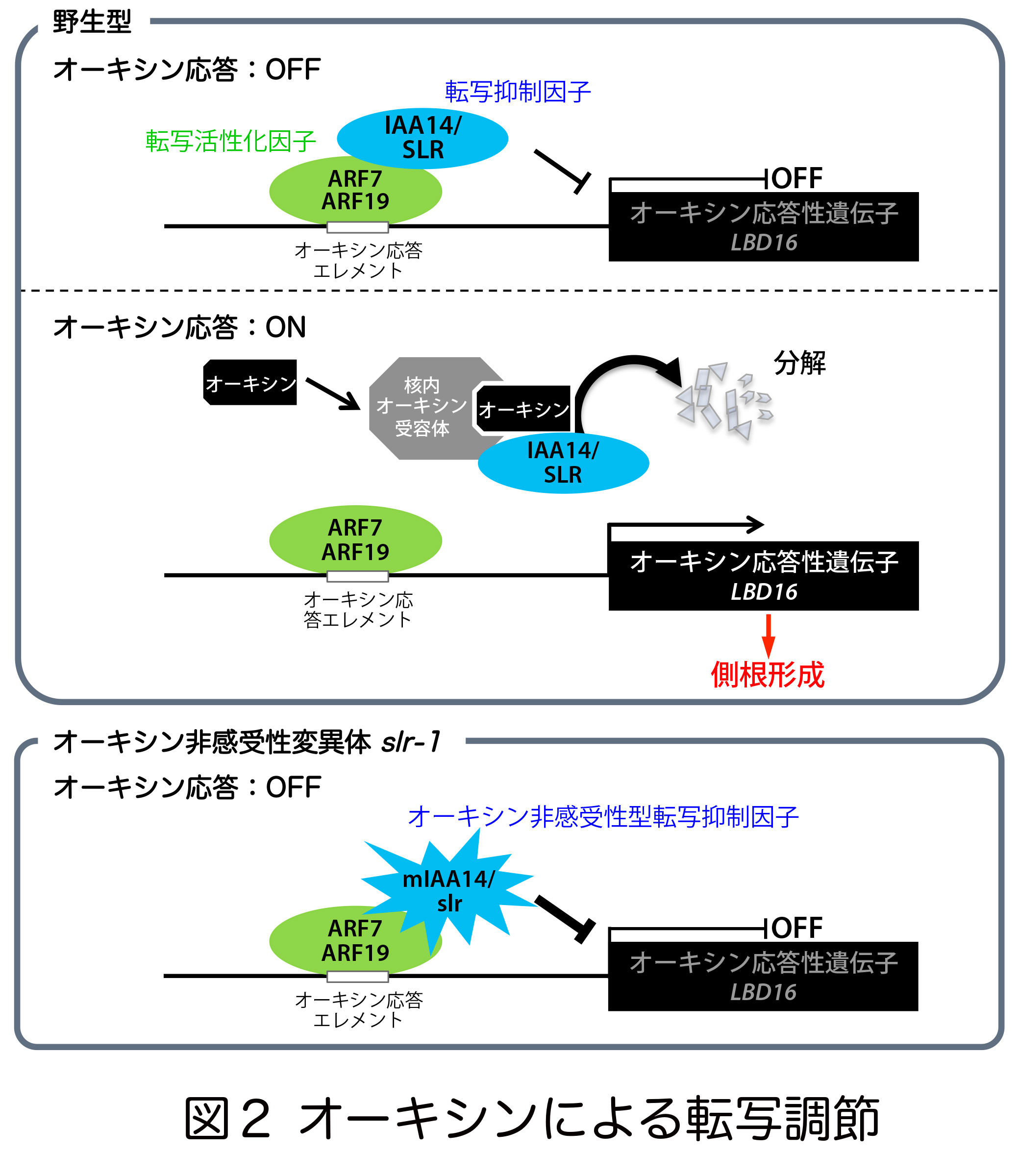 、ARF7とARF19がLBD16に代表される下流遺伝子の転写をONにします。逆に、オーキシンが存在しないかもしくはSLRに変異が入って安定化してしまうと、ARF7とARF19の転写活性化因子としての役割が阻害され、LBD16の転写がOFFになります(図2)。遺伝子の転写には転写装置(RNAポリメラーゼⅡ)の起動が必要なのですが、これらの転写調節因子が転写装置に動力を伝達する仕組みは謎でした。
、ARF7とARF19がLBD16に代表される下流遺伝子の転写をONにします。逆に、オーキシンが存在しないかもしくはSLRに変異が入って安定化してしまうと、ARF7とARF19の転写活性化因子としての役割が阻害され、LBD16の転写がOFFになります(図2)。遺伝子の転写には転写装置(RNAポリメラーゼⅡ)の起動が必要なのですが、これらの転写調節因子が転写装置に動力を伝達する仕組みは謎でした。
そこで古谷研究員らは、転写因子と転写装置をつなぐ転写メディエーター複合体に着目しました。転写メディエーター複合体は4つのモジュール(head/middle/tail/kinase)から構成され、head/middle/tailの3つのモジュールは転写装置を起動し、kinaseモジュール(CKM)は転写装置を停止します。
本研究では、IAA14/SLRが転写補助抑制因子TOPLESS(TPL)を介してkinaseモジュールと複合体を作り、下流遺伝子LBD16の転写をOFFにして発根を抑えることを明らかにしました。また、オーキシンによってIAA14/SLRが分解されると、転写メディエーター複合体からkinaseモジュールが外れ、LBD16の転写をONにして発根を促進していることが分かりました(図3)。 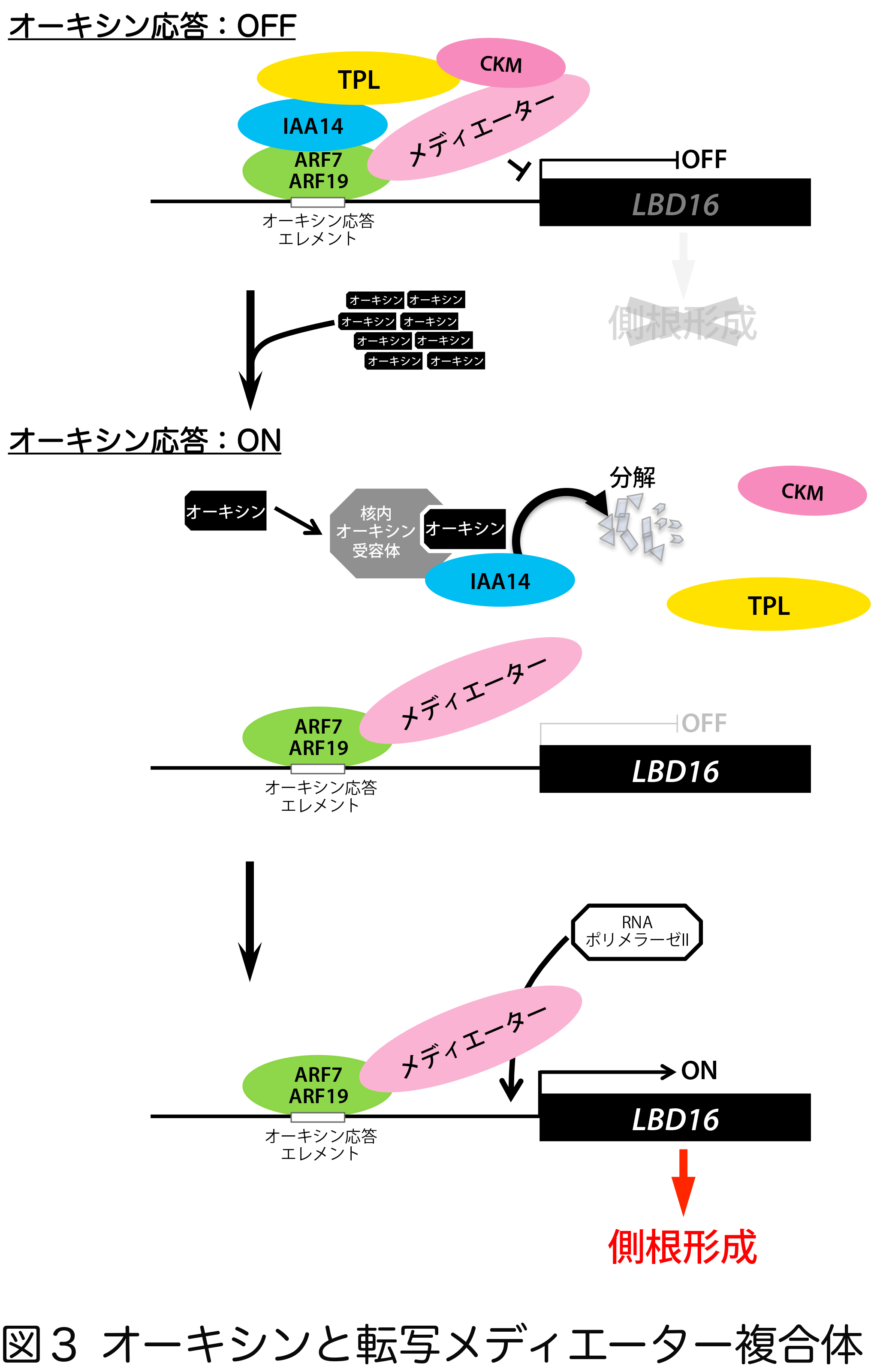
これらのことから、オーキシンによるkinaseモジュールの脱着が転写装置に動力を伝達し、下流遺伝子の転写のON・OFFを素早く切り替える仕組みが明らかとなりました。
【成果の意義】
本研究の成果により、オーキシンが発根を促進する仕組みが明らかとなりました。今後、オーキシンと同じように転写メディエーター複合体のかたちを変えることができる化学物質を発根促進剤として開発することで、希少または有用植物の挿木・挿苗による量産化技術の開発が期待されます。
【用語説明】
オーキシン:
植物ホルモンの一つ。発根のほか、地上部の葉や花の形成を促進する働きを持つ。
転写メディエーター複合体:
酵母から哺乳類、植物まで高度に保存されている。転写因子と転写装置を仲介する巨大なタンパク質複合体で、動物では癌との関連から研究が盛んに行われている。植物では最近、オーキシン以外の植物ホルモンや耐病性への関与が明らかになりつつある。
モジュール:
タンパク質複合体の一部を構成するひとまとまりの機能を持ったタンパク質の集合体。
【論文名】
掲載論文:Proc. Natl. Acad. Sci. U.S.A. 米国科学アカデミー紀要
論文タイトル:
Auxin-dependent compositional change in Mediator in
ARF7- and ARF19-mediated transcription
著者名:
Jun Ito, Hidehiro Fukaki, Makoto Onoda, Lin Li,
Chuanyou Li, Masao Tasaka, and Masahiko Furutani
【植物形態ダイナミクス】
研究室紹介ページ:http://bsw3.naist.jp/courses/courses106.html
研究室ホームページ:http://bsw3.naist.jp/keihatsu/
(2016年06月06日掲載)
この記事をシェア ![]()