Research 研究成果の紹介
バイオサイエンス研究科植物形態ダイナミクス研究室の古谷将彦助教・田坂昌生教授らが、花や葉を形作る分子メカニズムを解明(~器官の発生に必須なオーキシンの流れを生み出す仕組みを発見~ 自在に園芸植物をデザイン、増産にも期待)
バイオサイエンス研究科植物形態ダイナミクス研究室の古谷将彦助教、田坂昌生教授らは、花や葉を形作る分子メカニズムを解明した。
古谷将彦助教のコメント
今回の成果は、これまで明らかにされてこなかった、シンクとして機能するオーキ シンの流れを構築する分子機構の解明にあります。我々が単離・同定してきたMAB4遺伝子群がオーキシンシンクの確立に関与することを発見しました。また、MAB4遺伝子群がオーキシンによって発現誘導されることを見つけ、オーキシンの流れを構築するポジティブフィードバック機構が明らかになりました。今回の発見により、古くから提唱されている「オーキシン運河仮説」を分子レベルで説明できる可能性が出てきました。今後さらなる解析を行い、この仮説の実証に挑戦していきたいと思います。本研究は、当時大学院生であった中野泰一くんの大きな協力を得て行われました。この場を借りて感謝申し上げます。
シンの流れを構築する分子機構の解明にあります。我々が単離・同定してきたMAB4遺伝子群がオーキシンシンクの確立に関与することを発見しました。また、MAB4遺伝子群がオーキシンによって発現誘導されることを見つけ、オーキシンの流れを構築するポジティブフィードバック機構が明らかになりました。今回の発見により、古くから提唱されている「オーキシン運河仮説」を分子レベルで説明できる可能性が出てきました。今後さらなる解析を行い、この仮説の実証に挑戦していきたいと思います。本研究は、当時大学院生であった中野泰一くんの大きな協力を得て行われました。この場を借りて感謝申し上げます。
研究の概要
【概要】
奈良先端科学技術大学院大学 バイオサイエンス研究科 植物形態ダイナミクス研究室 古谷将彦助教、田坂昌生教授らは、植物が花や葉を作るさいに必要なホルモンであるオーキシンについて、それを植物体内で働く場所まで届かせるために「オーキシンの流れを生み出す」という基本的な仕組みを明らかにした。謎と言われた現象を分子遺伝学の手法により解明した。
植物ホルモンであるオーキシンは生合成された部位から機能する場所の方向に流れる形で輸送され、蓄積する。そして、茎の先端に存在する“芽”においてオーキシンが蓄積することにより、花や葉などの植物の器官が形成される。このような器官が形成される際のオーキシンの流れ方については明らかになりつつあるが、その流れを生み出す源の仕組みは謎であった。
古谷助教、田坂教授らは、独自に発見したオーキシンの流れを調節する遺伝子群が花の形成時期に機能することに着目。これらの遺伝子機能を失い、花を作らなくなった変異体を用いて分子遺伝学的解析を行った。その結果、この変異体ではオーキシンの流れが滞っていることを発見。このことから、これらの遺伝子群がオーキシンの流れそのものを生み出す源であることを明らかにした。さらに、これらの遺伝子群がオーキシンそのものによって誘導されることから、オーキシンの流れを増幅する仕組みが示された。
今後、オーキシンの流れを人工的に操作し花や葉の形や配置を自在に変えられる可能性があり、多くの大輪の花をつけるなど園芸植物の改変や農作物、植物バイオマスの増産が期待される。研究成果は平成26年1月6日付けで米国科学アカデミー紀要(Proceedings of the National Academy of Sciences of the United States of America)の電子版に掲載された。
【研究の背景】
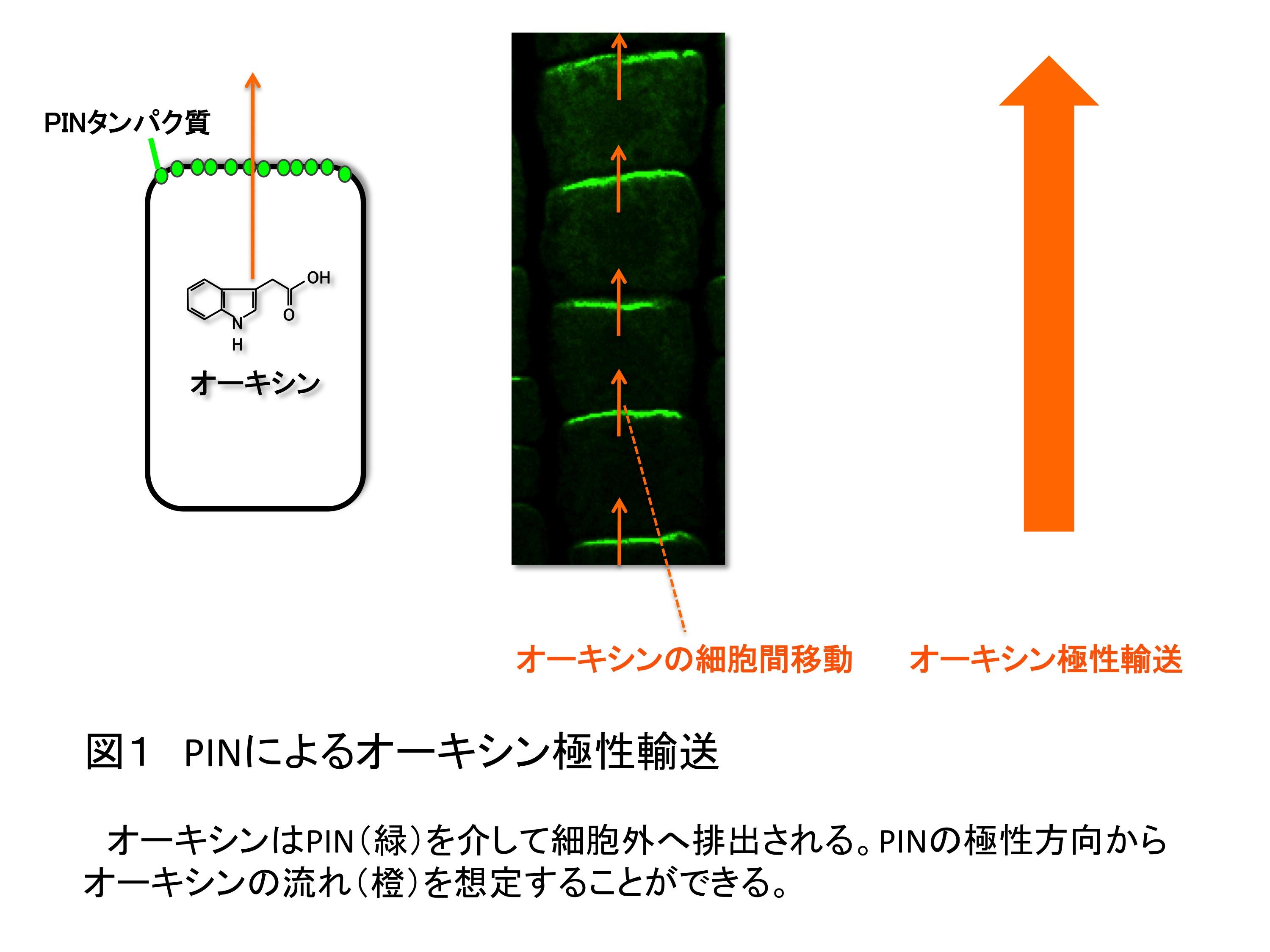
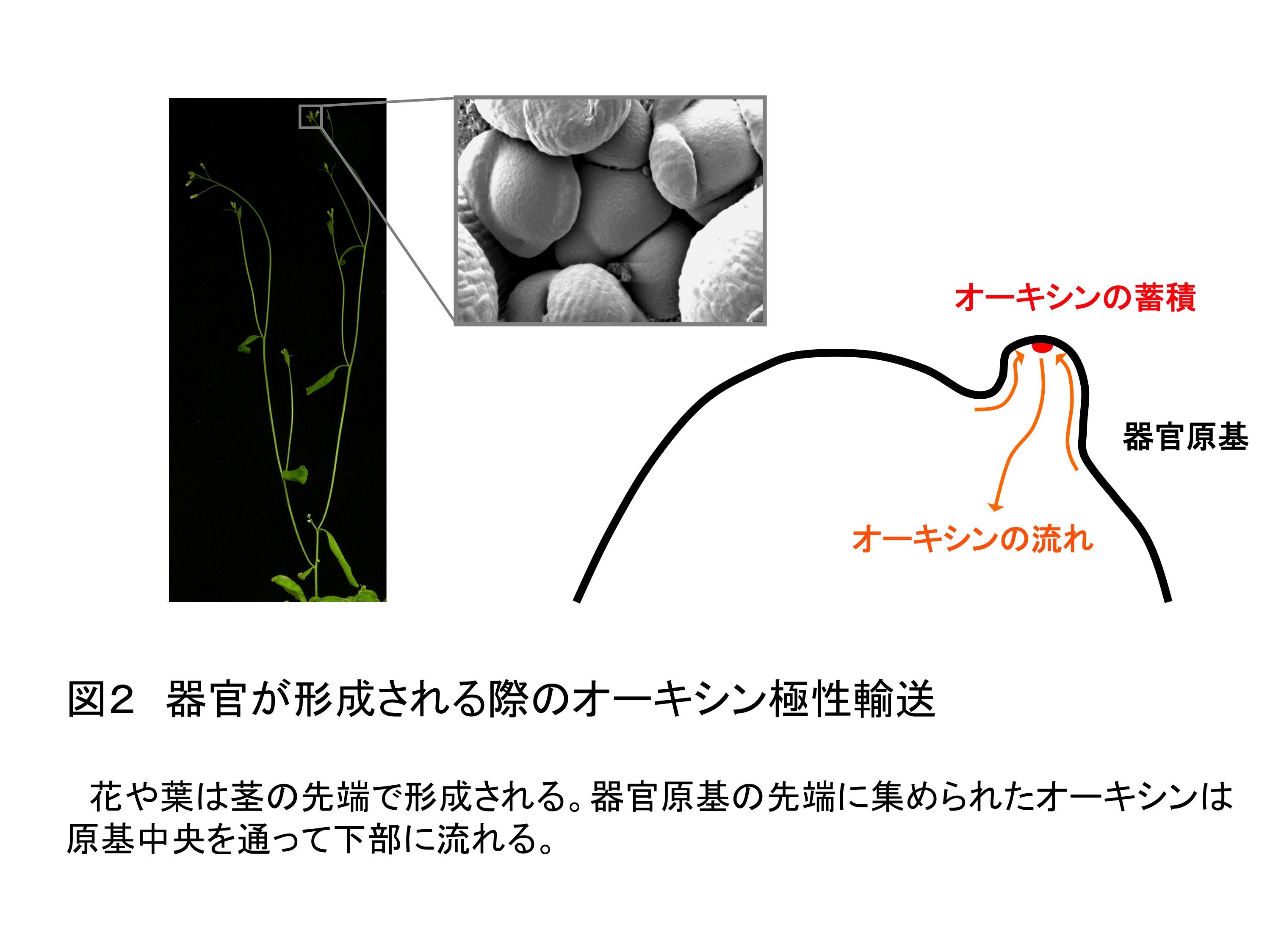
葉や花といった植物の器官は植物ホルモン・オーキシンに依存して形成される。オーキシンの蓄積部位に隆起が生じ、特定の器官に細胞が分化する器官原基へと発達するのである。このオーキシンの蓄積には、方向性をもったオーキシンの流れ「オーキシン極性輸送」が必須である。オーキシン極性輸送の仕組みは、まず、PIN-FORMED (PIN)と呼ばれるオーキシンを細胞外に排出する担い手となるタンパク質が個々の細胞の細胞膜上でずらっと並んで局在化し極性を持つ。次いで、つながった細胞の極性が一定の方向に揃うことで可能となる(図1)。器官が形成されるときには、一点にオーキシンを集中して蓄積できるような形にPINの配向がそろって変えられる(図2)。ところが、このPIN配向を決定する分子的な仕組みは明らかになっていなかった。
【結果】
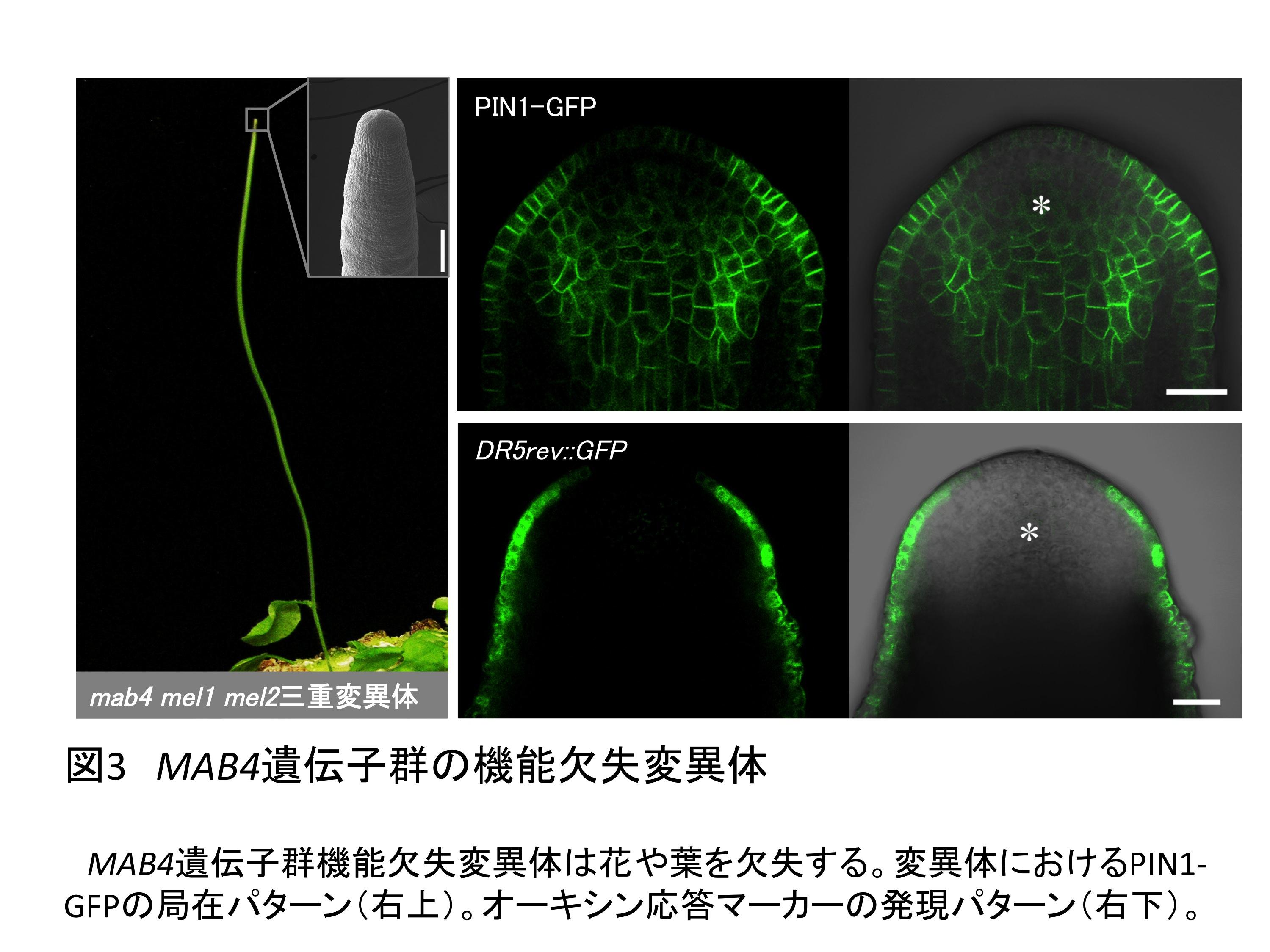
古谷助教、田坂教授らはPIN配向を確立する分子機構を明らかにするため、独自に同定した、オーキシンの流れを調節する役割があるMAB4遺伝子群の機能解析を行った。まず、MAB4遺伝子群の機能を欠失した変異体では花や葉を欠き、異常なPIN配向を示すとともにオーキシンの流れが停滞することが明らかになり、MAB4遺伝子群が器官形成時のオーキシンの流れを制御することが示された(図3)。さらに、MAB4遺伝子群はオーキシンの作用により誘導されて、機能が発現することが示され、このとき、オーキシンの流れがより増幅されるようにPIN配向が確立されるというポジティブフィードバック機構が明らかになった。
【本研究の意義】
オーキシンの流れを生み出す機構については、古くから多くの仮説が提唱されてきた。「オーキシン運河説」と呼ばれる仮説は、合流しながらしだいに大きな流れとなる川のように、オーキシンの流れが増強するように確立されるというもので、葉脈のパターンなど植物の発生現象を説明できるとして有名である。本研究で明らかにした分子機構はオーキシン運河説を説明しうる可能性があり、今後のさらなる解析から仮説を実証することができるかもしれない。また、本研究成果をもとにオーキシンの流れを人為的に操作することで葉や花などの器官の配置や形を変化させることが可能となり、園芸植物の品種改良や農作物、植物バイオマスの増産につながるであろう。
【補足説明】
●オーキシンとは
植物ホルモンの一種。天然のオーキシンとしてはインドール-3-酢酸が最も多く存在する。オーキシンは細胞の伸長、分裂、分化などを促進する作用があり、茎や根などの屈性反応(光や重力などの外部刺激に応答した成長運動)、根や葉など器官の発生、葉脈のパターン、胚発生などさまざまな生理・発生現象に関わる。このあまりに広範囲な作用そして植物成長への大きな貢献度からこれまで解析が困難であったが、近年ようやくオーキシンの受容体が同定され、そのシグナル伝達経路が明らかになりつつある。また、生合成経路についても徐々に明らかにされてきている。
●PINタンパク質とは
オーキシンを細胞外へと排出する膜タンパク質。細胞膜に極性を持って局在化し、オーキシンを細胞外のある方向へ排出する。細胞膜上のPINタンパク質は恒常的なエンドサイトーシスによって細胞内へと移行して、一部は液胞へと送られ分解される。分解を逃れた一部のPINタンパク質は再び細胞膜へと戻される。PINタンパク質の細胞内の分布はさまざまな環境変化、外部刺激、発生シグナルを受けて変化し、これが環境応答時や発生の場面におけるオーキシンの流れの変化の原動力になっている。
●MAB4遺伝子群とは
植物に特有のNPH3様タンパク質をコードするMACCHI-BOU 4遺伝子群の略称。MAB4遺伝子群はPINタンパク質の細胞内の分布を調節する因子として発見され、重力に応答した根の成長や子葉の発達に関わることが明らかになっている。コードするタンパク質は共通してBTB/POZドメインとNPH3ドメインをもつが、これらのドメインの機能は未解明である。また、PINタンパク質と同じように細胞膜上に極性を持って存在する。
(2014年02月04日掲載)
この記事をシェア ![]()