
2023.07.12
高湿度環境における植物と病原細菌の攻防
植物免疫学研究室・助教・安田 盛貴
- 要旨
- 降雨などがもたらす高湿度環境は、病原細菌による葉の病害を助長します。その要因として、葉内の病原細菌が引き起こす水浸漬現象が知られています。近年の研究から、病原細菌が水浸漬を誘導する仕組みの理解は進んでいますが、植物がどのように水浸漬を抑えているのか、その実態は不明な点が多く残されています。私たちはアブラナ科植物シロイヌナズナを用いて研究を進め、植物が有する水浸漬抵抗性の一端を突き止めました。本稿では、最初に病原細菌が水浸漬を誘導する仕組みを概説し、次に私たちが発見した植物のアクアポリンを介した水浸漬抵抗性を紹介します。
1.はじめに
細菌や糸状菌などの病原菌による植物の病害は、農作物や飼料の生産に甚大な被害をもたらし、時として人間の社会生活を大きく揺るがします。植物の発病は、感染した病原菌が宿主植物の有する抵抗性を打破できることに加え、それらを取り巻く温度、湿度、光などの環境条件に左右されます(図1)。植物は細胞膜上のパターン認識受容体(PRR: pattern recognition receptor)で病原菌由来、または感染時に産生される自己由来の分子パターンを認識し、防御応答を誘導することで病原菌の感染拡大を防いでいます。これはパターン誘導性免疫(PTI: pattern-triggered immunity)と呼ばれ、植物の基礎抵抗性を担います。近年の研究から、環境条件がPTIに与える影響とその仕組みが徐々に紐解かれてきています1)。また、病原菌は環境条件に依存して植物感染時に周囲の組織環境を増殖に有利な状態に変えることも分かってきました2)。私たちは、高湿度環境が植物と病原菌の相互作用に及ぼす影響を明らかにするため、アブラナ科植物シロイヌナズナと病原細菌Pseudomonas syringae pv. tomato(Pst)DC3000をモデルに研究を進めています。
図1.植物病害の要因となる3要素
宿主植物と病原菌の関係性に加えて、それらを取り巻く環境条件が鍵となる。
2.高湿度において葉内の病原細菌が誘導する水浸漬
気孔などの開口部から葉内に侵入した病原細菌Pst DC3000は(以降は病原細菌と表記)、細胞間の隙間(細胞間隙)に定着し、水と栄養を獲得することで増殖します。通常の栽培湿度(相対湿度約60%)と比べて、高湿度(相対湿度約95%)では病原細菌を接種した葉が著しい病徴を示します(図2)。その要因となるのが、葉内の病原細菌が細胞間隙に水を蓄積し増殖を促進する「水浸漬エリア」の誘導です(図2)3)。病原細菌はIII型分泌装置と呼ばれる注射針のような器官を使って、宿主シロイヌナズナの細胞内にエフェクターと呼ばれるタンパク質を送り込み水浸漬を誘導します。これまでに、HopM1およびAvrEと呼ばれる冗長的に働く2つのエフェクターによる水浸漬誘導の仕組みが報告されています3)4)5)。HopM1/AvrEは宿主シロイヌナズナの植物ホルモンABA(アブシシン酸)経路を活性化し、気孔閉鎖を誘導することで葉の保水性を高め、葉内の病原細菌による水浸漬を促します。HopM1/AvrEはABA経路の複数ステップを標的としており、ABA生合成、孔辺細胞(気孔を構成する細胞)への流入およびシグナル伝達に働きかけることが明らかにされています4)5)。このように、病原細菌はシロイヌナズナの乾燥ストレス適応を支えるABA経路を乗っ取り、それを葉内における水獲得に利用する巧妙な感染戦略を備えていることがわかってきました。

図2.湿度条件が葉の細菌病に与える影響
病原細菌Pst DC3000を葉内に接種したシロイヌナズナを通常湿度または高湿度で栽培し、1日後に水浸漬を、5日後に病徴を観察した。
3.水浸漬に対する植物の抵抗性
病原細菌が水浸漬を誘導する仕組みが明らかになる一方で、シロイヌナズナがどのように水浸漬の発生を防いでいるのか、その実態は不明なままでした。そのような状況のなか、私たちはシロイヌナズナの細胞膜アクアポリンPIP2;6による水浸漬抵抗性を見出しました(論文投稿準備中)(図3)。アクアポリンは生体膜に存在するチャネルタンパク質で、膜内外の浸透圧勾配に応じて水分子を輸送します。PIP2;6を欠損したシロイヌナズナ変異体(pip2;6)は、病原細菌のエフェクター非分泌株の接種時にも顕著な水浸漬を示します。PIP2;6のN末端に位置するスレオニン残基のリン酸化は、PIP2;6の水分子輸送活性を促進し、水浸漬の抑制に寄与します。興味深いことに、シロイヌナズナは高湿度に応じてロイシンリッチリピート型受容体様キナーゼBAK1を介してPIP2;6のリン酸化を誘導することが示されました。SERKファミリーに属するBAK1(および他のSERKs)は、PRRをはじめ多くのロイシンリッチリピート型受容体様キナーゼ/タンパク質と協調的に働き、細胞外リガンドを認識することから、高湿度に応じて産生される細胞外リガンドとその受容体の存在が示唆されます。PIP2;6が水浸漬抵抗性で機能を発揮するためには、BAK1を介したリン酸化に加えて、ARF-GEFタンパク質であるMIN7を介した細胞内輸送が必要となります(図3)。MIN7はエフェクターHopM1の標的のひとつであり、ユビキチン・プロテアソーム系による分解へと導かれます6)。シロイヌナズナ変異体と病原細菌変異株を用いた解析から、病原細菌はエフェクターHopM1/AvrEとエフェクターAvrPto/AvrPtoBを用いてMIN7経路とBAK1経路をそれぞれ阻害し、PIP2;6を介した水浸漬抵抗性を打破することが分かりました(図3)。植物は細胞内のヌクレオチド結合ロイシンリッチリピート受容体でエフェクターを直接または間接的に認識し、エフェクター誘導性免疫(ETI: effector-triggered immunity)と呼ばれる強固な防御応答を発動します。ETIの活性化は病原細菌による水浸漬を抑制することが示されており3)、これはpip2;6においても認められました。以上のことから、シロイヌナズナと病原細菌は、(1)高湿度によるPIP2;6依存的な水浸漬抵抗性の誘導、(2)エフェクターによるPIP2;6依存的な水浸漬抵抗性の打破、(3)エフェクター認識によるPIP2;6非依存的な水浸漬抵抗性の誘導、という攻防を繰り広げていると考えられます。
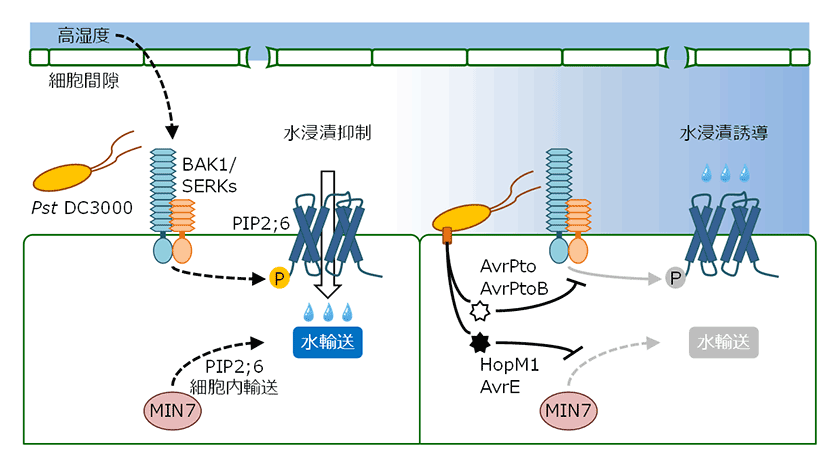
図3.シロイヌナズナの細胞膜アクアポリンPIP2;6による水浸漬抵抗性
高湿度に晒されたシロイヌナズナは、BAK1(および他のSERKs)によるリン酸化とMIN7による細胞内輸送でPIP2;6依存的な水浸漬抵抗性を誘導する。一方で、病原細菌Pst DC3000はエフェクターを用いてBAK1経路とMIN7経路を阻害し、PIP2;6依存的な水浸漬抵抗性を打破する。
4.おわりに
私たちの研究から、シロイヌナズナは高湿度に応じて水浸漬抵抗性を誘導することが示されました。本稿では触れませんでしたが、高湿度誘導性の遺伝子発現を示すABA代謝酵素を介した水浸漬抵抗性も見出しています。シロイヌナズナが高湿度に応答する仕組みの解明は、水浸漬抵抗性の更なる理解に加え、高湿度がPTIやETIにおよぼす影響を検証するうえで重要な課題です。今後は、シロイヌナズナの高湿度応答機構の解明に取り組み、植物が病害を助長する高湿度環境にどのように適応しているのか紐解くことで、そのような環境下でも頑健に病害を防除できる技術の開発に貢献していきたいと思います。
参考文献

著者
安田 盛貴 Researchmap
略歴
2014年北海道大学大学院生命科学院博士課程修了。
北海道大学博士研究員、奈良先端科学技術大学院大学博士研究員を経て、2022年より現職
- 研究内容:植物と病原細菌が高湿度に応答する仕組み
- 抱負:研究を楽しむ
- 関心ごと:健康維持
安田 盛貴 NAIST Edge BIO, 0014. (2023)