
2022.06.01
中枢神経系の発生における多様な細胞への分化と配置の制御
発生医科学研究室・助教・篠塚琢磨
- 要旨
- 多細胞生物のからだには様々な種類の細胞が存在し、それぞれの細胞が固有の機能を持っています。これら多様な細胞は組織内で一定の決まった順序で配置されています。このような細胞の多様性と配置の一定性を生み出すメカニズムとして、組織の一部の細胞において産生、分泌されるタンパク質(分泌性シグナル因子)が組織内で濃度勾配を形成し、その濃度を細胞が読み取ることで特定の細胞へと変化(分化)していく仕組みが知られています。本稿の前半では、細胞の多様性が生み出される一方で、それらが一定の順序で配置されるメカニズムの概念について解説します。後半では、実際に細胞の分化がシグナル因子により与えられる「位置情報」によって制御されている事例として、中枢神経系の多様な神経細胞が領域を形成するメカニズムを概説します。さらに、私たちが着目して研究を行っているグリア細胞の多様性と領域特異性について、今後の展望を含めて紹介します。
1.細胞の多様性と配置の一定性
胚発生期(受精卵が母体内や卵の中で成長する期間)には、未分化な細胞が細胞外部からもたらされる「情報」によって前駆細胞へと変化し、さらに特定の機能を持った細胞(成熟細胞)へと分化(成熟)していきます(図1A)。未分化な細胞が前駆細胞へと変化する際に、その細胞の運命決定に必須の情報を与えると考えられているのが、分泌性のシグナル因子(タンパク質)です。このシグナル因子は、胚発生期に形成される組織の中で、「細胞分化の司令塔」として働く特定の細胞集団(産生細胞)から分泌され、組織内で濃度勾配を形成します。一方、組織内の細胞(受容細胞)はこの濃度(情報)によって、自分が組織内のどの位置にいるのかを感知します(つまり「位置情報」を獲得する)ことになります。受容細胞にはシグナル因子に応答する閾値濃度が存在し、濃度に応じて遺伝子発現が変化することで、その細胞運命が決まります(図1B)。このように、単一の分泌因子が複数の細胞運命を規定し、それらを特定の順序で配置するシステムは、1969年、イギリスの生物学者ルイス・ウォルパート(Lewis Wolpert)博士によって「モルフォゲン説」として提唱されました1)。それ以来、この仮説は発生生物学における形態形成、細胞分化の重要な概念として考えられています。
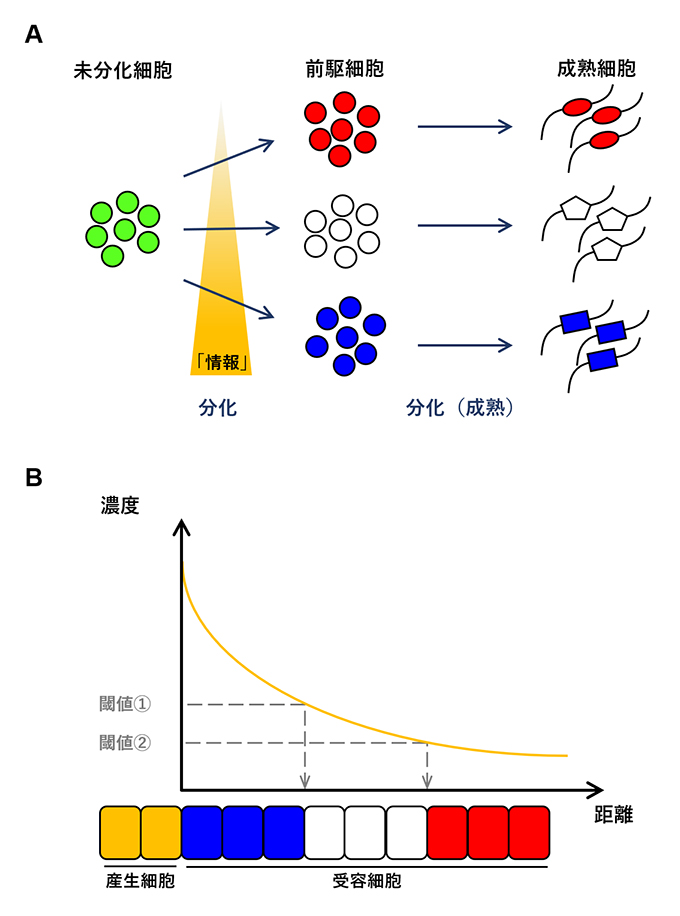
図1.
(A) 外部からの「情報」により未分化細胞が成熟細胞へと分化します。
(B) シグナル因子の濃度勾配により細胞の多様性と配置が決定されるメカニズムの概念「モルフォゲン説」。細胞の種類をフランス国旗のように3色に色分けすることができるので、「フランス国旗モデル」とも呼ばれています。
2.シグナル因子により生み出される神経細胞の多様性
脳・脊髄からなる中枢神経系は、個体外部からの刺激に対する反応、知性の表現に欠かせない制御システムです。神経細胞をはじめ、中枢神経系を構成する細胞は多様性に富んでおり、そのほとんどは胚発生期に生み出されます。これらの細胞の多様性が生み出されるメカニズムとして、胚発生期の中枢神経系は「モルフォゲン説」が適用される好例となっています。中枢神経系の発生において、胚の頭から尾に沿って、背中に神経管が形成されます。神経管からは脳、脊髄が作られますが、特に脊髄領域(体幹部)では、神経管内にShh(ソニック・ヘッジホッグ)、Wnt(ウィント)、BMP(ビーエムピー)といった分泌性シグナル因子が存在し、それらから与えられた「位置情報」により、神経前駆細胞の多様性と領域特異性が生まれます。
脊髄領域の神経管を背腹を含む面(横断面)で輪切りにすると、神経前駆細胞(神経に分化する細胞)が背側から腹側(背腹軸)に沿って、11種類の領域に分かれて並んでいることが観察されます。これらの神経前駆細胞は、それぞれ対応する神経細胞へと分化(成熟)していきます。この過程で、神経管の最も背側の領域(蓋板)からはWnt、BMPが、また最も腹側の領域(底板)からはShhが分泌され、これらの因子が神経管内に背腹軸に沿った濃度(活性)勾配を形成しています2),3)(図2)。神経管内にある神経前駆細胞は、この濃度のちがいを読み取り、濃度に応じて異なる細胞へと分化、つまり異なる機能を獲得します4)。たとえば、高濃度のShhが作用した神経前駆細胞はV3と呼ばれる介在神経細胞に、低濃度のShhが作用した神経前駆細胞は運動神経にそれぞれ分化します。
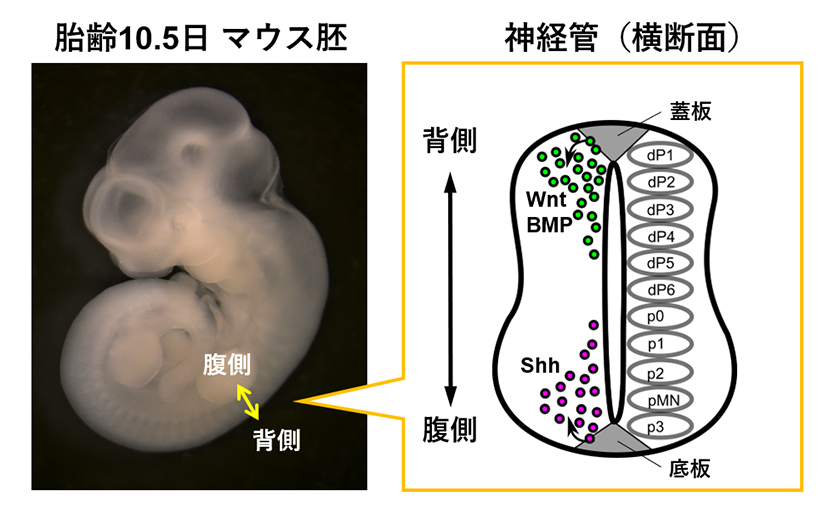
図2.胎齢10.5日のマウス胚の神経管横断面。神経前駆細胞が背腹軸に沿って11の領域(dP1-p3)に分かれています。
3.グリア細胞の機能多様性と領域特異的な分化
神経系を構成する細胞として、神経細胞のほかにグリア細胞の存在が知られています。グリア細胞は神経系の大部分を占める細胞であり、神経細胞の生存や機能をサポートするなど、神経系の中で「縁の下の力持ち」として働くと考えられてきました。これまで多くの研究は神経細胞に焦点が当てられていましたが、近年、グリア細胞の機能と多様性が注目されてきています。
グリア細胞の起源も、神経細胞と同じく神経前駆細胞です。脊髄領域の神経管では神経前駆細胞からのニューロンの分化は発生の中期頃までに完了し、その後はグリア細胞が生じます5)。グリア細胞はオリゴデンドロサイト、アストロサイトなどに分類され、細胞ごとに異なる機能を持っています。そして、これらの細胞の多様性も、Shh、Wnt、BMPといったシグナル因子が作り出す濃度勾配によって生み出されると考えられています。たとえば、大部分のオリゴデンドロサイトは、Shhが作用した神経管腹側の特定の神経前駆細胞から産生されます6)。また、作用したShhの濃度に応じて、VA1、VA2、VA3と呼ばれるアストロサイトが神経前駆細胞からそれぞれ領域特異的に産生されることが示されています7)。しかし、グリア細胞の多様性を生み出す分子メカニズムの研究は、神経細胞と比較して発展途上であり今後の課題として残されています。
4.In vitro 神経分化システムを用いたグリア細胞の多様性の解析
上述したように、グリア細胞(特にアストロサイト)にも多様性が存在し、それぞれ発現する遺伝子や機能が異なることが予想されますが、これらのグリア細胞の種類を定義するためのマーカー遺伝子は神経細胞と比べてほとんど同定されていません。
そこで、私たちはグリア細胞の多様性を生み出す分子メカニズムを明らかにするため、マウス胚性幹細胞(ES細胞)を用いたin vitroでの神経分化システム8),9)を応用した解析を行っています。ES細胞の分化は生体内ではなくシャーレ上で誘導するため、分化途上にある細胞に対して任意のシグナル因子を自由な濃度で添加することができるという利点があります。すなわち、対象となる細胞のインプット(シグナル因子)とアウトプット(遺伝子発現・細胞機能)を対応づけることが容易です。また、細胞の分化状態の変遷をリアルタイムに追跡することも可能です。これまでに私たちは、ShhやWntといったシグナル因子を様々な濃度で組み合わせることにより、ES細胞から、特定の神経前駆細胞のみならず、グリア細胞へと分化させることに成功しています(図3)。このようにして作成されたグリア細胞は、生体内においてシグナル因子によって与えられた「位置情報」から領域特異的に分化したグリア細胞と対応することが期待されます。今後は、これらのグリア細胞の解析を進め、特定の種類のグリア細胞を規定するマーカー遺伝子を同定し、グリア細胞の機能多様性と領域特異性の関係を明らかにすることを目指しています。
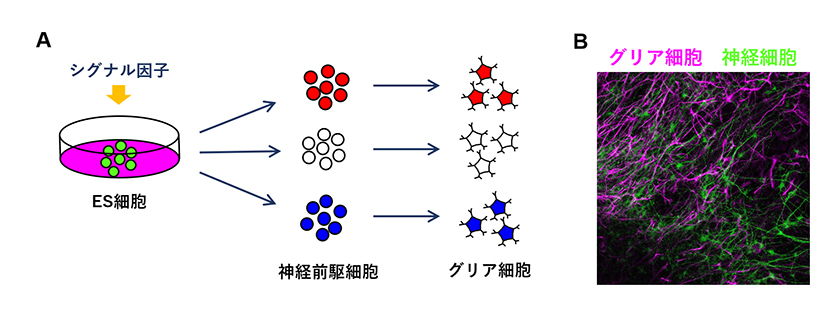
図3.
(A) ES細胞から特定のグリア細胞に分化させます。
(B) ES細胞から分化させたニューロン(緑)およびアストロサイト(マゼンタ)。
5.おわりに
本稿では、モルフォゲン説にはじまり、発生過程における神経細胞およびグリア細胞の多様性を生み出す分化メカニズムについて論じましたが、中枢神経系においてグリア細胞は発生期のみならず成体でも産生され、個体や神経系の恒常性の維持に深く関与しています。たとえば、上衣細胞といった一部のグリア細胞は神経幹細胞として知られるとともに、神経の損傷に応答して反応性アストロサイトへと変化したり、グリア瘢痕(はんこん)を形成したりすることで神経の再生に関与する性質を持っています10)。しかし、そのとき産生されるグリア細胞の種類や分化メカニズムに関する知見は限定的です。私たちの研究を通して、グリア細胞の分化メカニズムの理解が深まれば、神経損傷に対する生体反応や、神経再生・治癒を促すメカニズムの解明につながり、将来的には再生医療の発展にも貢献できると期待しています。
参考文献
- Kerszberg and Wolpert, Cell(2007).
- Shinozuka and Takada, J. Dev. Biol(2021).
- Yamada et al., Cell(1993).
- Jessell, Nat. Rev. Genet.(2000).
- Miller and Gauthier, Neuron(2007).
- Richardson et al., Nat. Rev. Neurosci.(2006).
- Hochstim et al., Cell(2008).
- Gouti et al.,PLoS Biol.(2014).
- Kutejova et al.,Dev. Cell(2016).
- Silver and Miller, Nat. Rev. Neurosci.(2004).

著者
篠塚 琢磨 Researchmap
略歴
2019年,総合研究大学院大学生命科学研究科にて博士号取得。
同年,基礎生物学研究所NIBBリサーチフェロー(博士研究員)。
2020年,同研究所特任助教を経て,2022年より現職。
- 研究内容:グリア細胞の多様性。思うようにグリア細胞に分化せず、頭や尻尾、ときには神経系ですらなく筋肉に分化してしまうES細胞と日々格闘中。
- 抱負:健康第一
- 関心ごと:陸上、登山、キャンプ、自転車
篠塚 琢磨 NAIST Edge BIO, 0001. (2022)